 | |
МЕНЮ
- Главная
- Языкознание филология
- Финансовые науки
- Управленческие науки
- Товароведение
- Технология
- Теплотехника
- Теория организации
- Теория государства и права
- Таможенная система
- Схемотехника
- Строительство
- Страхование
- Статистика
- Религия и мифология
- Психология и педагогика
- Промышленность производство
- Медицинские науки
- Медицина
- Краеведение и этнография
- Компьютерные науки
- История
- Искусство и культура
- Информатика
- Инвестиции
- Издательское дело и полиграфия
- Зоология
- Журналистика
- Естествознание
- Деньги и кредит
- Делопроизводство
- Гражданское право и процесс
- Государство и право
- Геополитика
- Геология
- Геодезия
- География
- Военная кафедра
- Ветеринария
- Валютные отношения
- Бухгалтерский учет и аудит
- Ботаника и сельское хоз-во
- Биржевое дело
- Биология и химия
- Биология
- Безопасность жизнедеятельности
- Банковское дело
- Астрономия
- Астрология
- Архитектура
- Арбитражный процесс
- Административное право
- Авиация и космонавтика
- Карта сайта
Книга: Мононуклеарный онкогенез
5. Инициация костномозгового мононуклеара
Костный мозг является одновременно органом кроветворения и центральным органом иммунной системы. Общая масса костного мозга у взрослого человека примерно 2,5–3,0 кг (4,5–4,7% от массы тела). Около половины составляет красный костный мозг, располагающийся в ячейках губчатого вещества плоских и коротких костей, эпифизов длинных (трубчатых) костей. Остальное – желтый костный мозг, выполняющий костномозговые полости диафизов длинных (трубчатых) костей.
В состав костного мозга входят стволовые клетки, из которых формируются основные клеточные элементы крови, клеточные элементы скелетных тканей, клеточные элементы сосудов. Чем моложе человек, тем активнее стволовые клетки костного мозга.
1. ГЕМОПОЭЗ
Гемопоэз (от греч. háima – кровь и póiēsis – сотворение) или кроветворение – процесс образования, развития и созревания клеток крови у животных и человека. Все клетки крови происходят из единой родоначальной клетки – полипотентной стволовой кроветворной клетки в эмбриогенезе и после рождения, они обладают способностью к дифференцировке во все без исключения клетки крови и возможностью участия в образовании других тканей организма. Различают эмбриональный и постэмбриональный гемопоэз.
ЭМБРИОНАЛЬНЫЙ
Эмбриональный гемопоэз представляет собой процесс развития крови, как ткани в эмбриональном периоде развития человека в результате последовательно сменяющихся трех основных этапов:
1. Первый этап – мезобластический: у человека начинается в конце 2-й – в начале 3-й недели, протекает по 9-ю неделю развития зародыша и характеризуется появлением первой генерации стволовых клеток крови. При этом клетки крови появляются во внезародышевых органах: мезенхиме стенки желточного мешка, мезенхиме хориона и стебля.
Кроветворение в стенке желточного мешка: в мезенхиме обособляются зачатки сосудистой крови или кровяные островки. В них клетки округляются, теряют отростки и преобразуются в стволовые клетки крови. Клетки, ограничивающие кровяные островки, уплощаются, соединяются между собой и образуют эндотелиальную выстилку будущего сосуда. Часть стволовых клеток дифференцируется в первичные клетки крови (бласты).
В поддержании плюрипотентности эмбриональных стволовых клеток млекопитающих, большое значение играют фибробласты в качестве подложки-фидера (от англ. feed – кормление, питание).
2. Второй этап – гепатолиенальный: начинается с 5–6-й недели развития плода, когда печень становится основным органом гемопоэза, в ней образуется вторая генерация стволовых клеток крови. Кроветворение в печени достигает максимума через 5 месяцев и завершается перед рождением. Стволовые клетки крови печени заселяют тимус, селезенку и лимфатические узлы.
3. Третий этап – медуллярный (костномозговой): начинается с 10-й недели и постепенно нарастает к рождению, характеризуется появлением третьей генерации стволовых клеток крови в красном костном мозге. После рождения красный костный мозг становится центральным органом гемопоэза.
Необходимо отметить, что все органы, в которых осуществляется кроветворение: лимфатические узлы, печень, селезенка, вилочковая железа и костный мозг, обладают определенными анатомо-физиологическими особенностями и имеют общие черты строения:
– строму составляет ретикулярная ткань, паренхиму – кроветворные клетки;
– богаты элементами, относящимися к системе Мононуклеарных фагоцитов;
– имеют капилляры синусоидного типа.
ПОСТЭМБРИОНАЛЬНЫЙ
Постэмбриональный гемопоэз представляет собой процесс физиологической регенерации крови, компенсирующий физиологическое разрушение дифференцированных клеток. Нормальное кроветворение осуществляется одновременно многими клонами – поликлонально и все клетки крови имеют костномозговое происхождение.
Структура костного мозга
Костный мозг обладает определенными анатомо-физиологическими особенностями. Его строму составляет ретикулярная ткань, паренхиму – кроветворные клетки. У человека красный костный мозг является высоко васкуляризированным органом, сообщается с кровотоком посредством капиллярной сети. Различают два типа капилляров: питающие (обычные) и функциональные (синусоиды), которые впадают в общий ствол – центральную вену.
Синусоиды располагаются радиарно, между ними находится гемопоэтическая ткань. В синусах между эндотелиальными клетками имеются поры, связывающие ткань кроветворных органов с кровеносным руслом. Такое строение обеспечивает транспорт клеток в кровь, а также поступление из крови в кроветворные органы гуморальных факторов (гемопоэтинов), которые наряду с нервной системой оказывают влияние на кроветворение. Будучи гистогенетически единой, кроветворная система в своем функционировании характеризуется определенной независимостью поведения отдельных ростков кроветворения.
Интенсивность гемопоэза
Владимирская Е.Б. (2001): у мужчин массой тела 70 кг, в сутки образуется 1 х 10 в 11 степени лейкоцитов и 2 х 10 в 11 степени эритроцитов, что составляет клеточную массу в 300 грамм (100 + 200 грамм соответственно) в сутки, в месяц образуется 9 кг клеток, в год – около 100 кг. За 70 лет жизни клеточная продукция нормального кроветворения очень велика и составляет порядка 7 тонн клеток крови.
Кроветворение в красном костном мозге
Кроветворение в красном костном мозге происходит островками, в которых группируются клетки по росткам гемопоэза. Предшественники и развивающиеся кроветворные клетки расположены следующим образом: в центре – делящиеся и незрелые клетки, на периферии (около стенок синусоидов) – более зрелые клетки.
Из полипотентной стволовой кроветворной клетки в красном костном мозге формируются все форменные элементы крови, развитие которых происходит экстраваскулярно. Часть их сохраняется в костном мозге в недифференцированном состоянии. Они могут расселяться по другим органам и тканям и являться источником развития клеток крови и соединительной ткани. Полипотентные стволовые кроветворные клетки относятся к самоподдерживающейся популяции клеток, и они редко делятся.
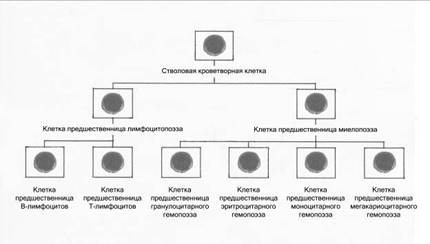
Рис. 6. Обобщенная схема первых трех классов кроветворения:
1 класс – полипотентная стволовая кроветворная клетка.
П класс – полипотентные клетки-предшественницы лимфоцито- и миелопоэза.
Ш класс – унипотентные клетки-предшественницы Т и В-лимфоцитов; би- и унипотентные клетки-предшественницы гранулоцитарного, эритроцитарного, моноцитарного и мегакариоцитарного гемопоэза.
Все стадии костномозгового кроветворения составляют шесть основных классов:
I класс – полипотентная стволовая кроветворная клетка – общая родоначальная клетка. Каждая полипотентная стволовая кроветворная клетка образует одну колонию и называется колониеобразующей единицей (КОЕ). Исследование клеточного состава колоний позволило выявить две линии их дифференцировки.
II класс – одна линия дает начало полипотентной клетке-предшественнице – родоначальнице гранулоцитарного, эритроцитарного, моноцитарного и мегакариоцитарного ростка гемопоэза – родоначальнице миелопоэза (КОЕ-ГЭММег). Вторая линия дает начало полипотентной клетке-предшественнице – родоначальнице лимфоцитопоэза (КОЕ-Л).
III класс – из полипотентных клеток дифференцируются би- и унипотентные родоначальные клетки. Методом колониеобразования определены би- и унипотентные клетки-предшественницы родоначальницы моноцитов (КОЕ-М), гранулоцитов и моноцитов (КОЕ-ГМ), нейтрофильных гранулоцитов (КОЕ-Гн), эозинофилов (КОЕ-Эо), базофилов (КОЕ-Базо), эритроцитов (БОЕ-Э и КОЕ-Э), мегакариоцитов (КОЕ-Мег), мегакариоцитов и эритроцитов (КОЕ-МегЭ). И унипотентные родоначальные клетки-предшественницы для Т- и В-лимфоцитов.
IV класс – из би- и унипотентных родоначальных клеток-предшественниц образуются бластные клетки различных линий гемопоэза, идентифицируемые при анализе миелограммы (эритробласт, мегакариобласт и т.д.);
V класс – созревающие клетки различных линий гемопоэза.
VI класс – зрелые клетки различных линий гемопоэза.
Все клеточные элементы, входящие в первые 3 класса и онтогенетически связанные, морфологически – не дифференцируемы (Рис. 6). Если даже обращается внимание на особенности в структуре ядра и цитоплазмы, то при изучении гематологических препаратов, их относят к лимфоидным элементам и бластным клеткам. Морфологически распознаваемые – это клетки 1У, У и У1 классов.
В процессе дифференцировки морфологически распознаваемые клетки эритроцитарного ряда претерпевают 5–6 митозов, гранулоцитарные клетки – 4 митоза, при моноцитопоэзе от монобласта до макрофага происходит 7–8 митозов.
Образующиеся в красном костном мозге клетки по мере созревания равномерно поступают в кровеносное русло, при этом время циркуляции их постоянно: эритроциты 100–120 суток и погибают, тромбоциты около 7–10 суток и погибают, нейтрофилы менее 6–8 часов и погибают, моноциты 1,5–4,5 суток и выходят в ткани.
МИКРООКРУЖЕНИЕ
Миелоидная и лимфоидная ткани относятся к тканям внутренней среды и являются разновидностями соединительной ткани. В них представлены две основные клеточные линии – клетки ретикулярной ткани и гемопоэтические клетки, т.е. для них характерно наличие стромальных и гемопоэтических элементов, образующих единое функциональное целое.
Ретикулярные, а также жировые, тучные и остеогенные клетки вместе с межклеточным веществом формируют микроокружение для гемопоэтических элементов. Структуры микроокружения и гемопоэтические клетки функционируют в неразрывной связи друг с другом. Микроокружение оказывает воздействие на дифференцировку клеток крови при контакте с их рецепторами или путем выделения специфических факторов.
РЕГУЛЯЦИЯ
Стволовые кроветворные клетки в стадии созревания находятся под строгим регулирующим контролем, механизм которого достаточно не изучен. В регуляции процессов пролиферации и дифференцировки гемопоэтических клеток большую роль играют:
1. Стромальное микроокружение:
– клеточный компонент: фибробласты, жировые клетки, макрофаги, остеобласты, эндотелиальные клетки;
– внеклеточный (экстрацеллюлярный) матрикс, который составляют продукты секреции стромальные клеток: коллаген, фибронектин, ламинин, гликозаминогликаны и другие белковые компоненты.
2. Факторы роста – обеспечивают пролиферацию и дифференцировку стволовых кроветворных клеток, и последующие стадии их развития. Факторы роста включают:
– колониестимулирующие факторы (КСФ) – стимулируют гемопоэз. Среди них наиболее изучены факторы, стимулирующие развитие гранулоцитов и макрофагов (ГМ-КСФ, Г-КСФ, М-КСФ);
– ингибирующие факторы – тормозят гемопоэз. Выделен ингибирующий лейкемию фактор (ЛИФ), который тормозит пролиферацию и дифференцировку моноцитов-макрофагов;
– интерлейкины.
3. Факторы транскрипции – влияют на экспрессию генов, определяют направление дифференцировки гемопоэтических клеток (поэтины).
4. Витамин В12 – необходим для стимуляции пролиферации и дифференцировки гемопоэтических клеток.
Таким образом, кроветворение в красном костном мозге – это единственная локализация ткани в организме человека, где в эмбриональном и постэмбриональном периодах жизни сконцентрировано большое количество клеток различной потентности и происходит интенсивная пролиферация.
2. МОНОНУКЛЕАРНАЯ ФРАКЦИЯ В СИСТЕМЕ КРОВИ
Мононуклеарная фракция в системе крови – это клетки, выделенные из костного мозга или периферической крови посредством отделения от эритроцитов, тромбоцитов и гранулоцитов на градиенте плотности.
МОНОЦИТОПОЭЗ
Моноцитопоэз берет свое начала от полипотентной клетки-предшественницы родоначальницы миелопоэзе с последующим развитием в Моноцитарный росток (П класс), продолжается в унипотентную клетку-предшественницу родоначальницу Моноцитов (Ш класс). Однако клеточные элементы, входящие во П и Ш классы морфологически не дифференцируемы. Морфологическое распознавание начинается с бластной клетки Моноцитарного ростка (1У класс) – монобласт, который через стадию Промоноцит, превращается в Моноцит.
В отличие от клеток других линий, цикл созревания которых заканчивается в красном костном мозге, клетки Моноцитарного ростка окончательно созревают только в тканях, где Промоноцит и Моноцит трансформируются в Макрофаг.
Морфология клеток:
По мере дифференцировки монобласта в промоноцит и моноцит клетка претерпевает ряд морфологических и функциональных изменений (Рис. 7):
В красном костном мозге
1. Монобласт – диаметром 12–20 мкм. В норме его трудно отличить от миелобласта или недифференцируемого бласта, а также не всегда можно отличить от лимфобласта. Только отмеченные очертания ядра и более широкая светло-базофильная цитоплазма могут указать на развитие этого «бласта» в сторону моноцитарной клетки. Ядро нежной структуры содержит 1–2 нуклеолы голубоватого цвета. Цитоплазма голубого цвета, в ней могут присутствовать пылевидные азурофильные гранулы.
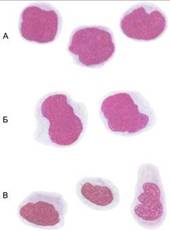
Рис. 7. Моноцитарный ряд клеток: А – монобласты, Б – промоноциты, В-моноциты.
2. Промоноцит – диаметром 15–20 мкм. В норме имеет ядро промиелоцита бобовидной формы, светло-фиолетового цвета. Хроматин нежный, крупносетчатый. В ядре 1–2 нуклеолы. Цитоплазма серо-голубого, дымчатого цвета с мелкой азурофильной зернистостью.
Промоноцит, являясь клеткой-предшественницей Моноцита, проходит 2 последовательных цикла деления до превращения в Моноцит, продолжительность митотического цикла составляет 30 часов. Промоноцит способен к пиноцитозу и фагоцитозу, хотя в меньшей степени, чем Моноцит и Макрофаг.
3. Моноцит – диаметром 16–18 мкм, различных морфологических вариаций по характеру и интенсивности окраски ядра и цитоплазмы. Ядра могут приближаться к округлым, бобовидным формам. Моноцит с более нежной структурой ядра и наличием ядрышек (или их остатков) можно отнести к Промоноциту. Цитоплазма сероватого или бледно-голубого цвета, в ней могут присутствовать многочисленные пылевидные азурофильные гранулы.
Дифференцировка Монобласта в Моноцит происходит в красном костном мозге в течение 5 дней. Моноцит в костном мозге находится в среднем 3 суток (минимальное время пребывания 9 часов), затем делится и, не образуя костномозгового резерва, выходит в периферическую кровь.
В крови
Моноцит – наиболее крупная клетка крови, здесь он созревает, ядро становится из круглого сначала бобовидным, затем лапчатым, меняется структура хроматина. В периферической крови обнаружен различный уровень дифференцировки Моноцитов, причем у здоровых людей преобладают более зрелые Моноциты. В незрелом Моноците есть остатки нуклеол, меняются ферменты в цитоплазме.
В крови Моноциты распределяются на пристеночные и циркулирующие пулы, обменивающиеся между собой, количественные соотношения которых могут меняться. У человека циркулирующий пул Моноцитов в норме 18х10 в 6 степени клеток/кг массы тела, а маргинальный пул, который в данный момент не принимает участия в циркуляции, примыкая к внутренней стенке микрососуда, в 3,5 раза больше (63х10 в 6 степени клеток/кг). В целом общий пул Моноцитов периферической крови составляют от 1 до 10% всех лейкоцитов (80–600 х 10 9/л).
Моноциты циркулируют в крови от 36 до 104 часов (1,5 – 4.5 суток) и затем покидают ее по стохастическому (целевому) принципу, взаимодействуя со специализированными адгезивными молекулами на эндотелиальных клетках. Миграция Мононуклеара из сосудистого русла в очаг воспаления происходит через участки микроциркуляторного русла с эндотелием второго типа – это посткапилляры и венулы.
В ткани
Van Furth R. (1988): после попадания в ткань Мононуклеары крови превращаются в Макрофаги, которые в свою очередь адаптируются к микросреде их будущего обитания. Разнородность популяции Макрофагов «по горизонтали» – микросреда, в которой они функционируют.
Выйдя из кровеносного русла, Моноцит становится тканевым и больше не способен вернуться в циркуляцию. Тканевой Моноцит трансформируется в органо- и тканеспецифический Макрофаг согласно следующими стадиям перехода: макрофагальный бласт, промакрофаг, макрофаг. Макрофаг может образоваться из стволовой кроветворной клетки и из Промоноцита согласно тем же стадиям перехода.
4. Макрофаг (от греч. makros – большой, fagos – пожирающий) – гетерогенная специализированная клеточная популяция защитной системы организма (Рис. 8). Диаметром 15–80 мкм, форма клетки неправильная, ядро овальной или продолговатой формы. Продолжительность жизни исчисляется месяцами и годами.
Возможно образование Макрофагов: в соединительной ткани (гистиоциты), легких (альвеолярные), печени (купферовские клетки), селезенке, лимфатических узлах, костной ткани (остекласты), нервной ткани (микроглиальные клетки), коже (клетки Лангерганса), в плевральном выпоте и асците и т.д.
Макрофаги принимают активное участие: в неспецифической защите от патогенных микроорганизмов; в процессах репарации; инициации специфического иммунного ответа; в метаболизме липидов и железа; регуляции кроветворения; гемостазе; в секреции цитокинов и других биологически активных веществ, регулирующих пролиферацию, дифференцировку и функциональную активность различных клеток.
По разнообразию фенотипических признаков Макрофагов можно судить о возможностях при трансформации Промоноцита и Моноцита и влиянии микроокружения. Макрофаг в сравнении с Промоноцитом и Моноцитом, по своим функциональным возможностям значительно превалирует. Значит, в процессе трансформации Макрофаг приобретает возможности, которые отсутствовали у Промоноцита и Моноцита.
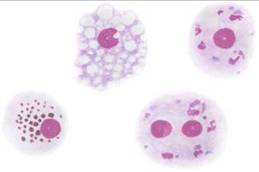
Рис. 8. Макрофаги
Промоноциты, Моноциты и Макрофаги человека в патологических условиях, способны к пролиферации в ограниченных масштабах. Промоноцит из этого ряда наиболее пролиферирующая клетка и может, минуя стадию Моноцита, в тканях трансформироваться в Макрофаг.
СТИМУЛЯЦИЯ
Учитывая, что Мононуклеары относятся к иммунокомпетентной системе, вполне законно ожидать изменение их количественного и качественного состояния при различных патологических процессах. В частности это наглядно проявляется при хроническом воспалении, когда число Моноцитов увеличивается. Это увеличение отмечается в красном костном мозге, периферической крови, селезенке, лимфатических узлах, перитонеальных полостях. При этом в периферической крови обнаружен различный уровень дифференцировки Моноцитов, а также могут появляться и преобладать менее зрелые клетки – Промоноциты.
В ускоренном вариант пролиферации костномозговых Мононуклеаров получается: минимум 9 часов в красном костном мозге и 12 часов в кровеносном русле, итого 21 час. Для созревания Моноцита из Промоноцита необходимо два деления по 30 часов, т.е. при срочной потребности в Макрофагах, с учетом отсутствия костномозгового резерва, в кровеносное русло выходят Промоноциты и не зрелые Моноциты.
В процессе дифференцировки морфологически распознаваемые клетки при Моноцитопоэзе претерпевают от Монобласта до Макрофага 7–8 митозов. При воспалительных процессах количество Макрофагов и их активность особенно возрастают. Воспалительный инфильтрат может поддерживаться лишь в случае постоянного обновления Мононуклеарных фагоцитов в очаге за счет Моноцитов крови. В свою очередь пул Моноцитов восстанавливается из красного костного мозга. Следовательно, через Мононуклеарные фагоциты очаг хронического воспаления приобретает постоянные связи с костным мозгом, с центром генерации свежих Моноцитов. Очаг будет прогрессировать при избыточном Моноцитопоэзе и наоборот. В то же время в стимуляции Моноцитопоэза самое деятельное участие принимают активированные Макрофаги.
Таким образом, Промоноцит и Моноцит в системе кроветворения являются единственными промежуточными упрощенными универсальными клетками, которые в органах и тканях трансформируются в органо- и тканеспецифические Макрофаги.
3. МОНОНУКЛЕАР – КЛЕТКА-ПРЕДШЕСТВЕННИЦА ЗЛОКАЧЕСТВЕННОЙ КЛЕТКИ
Клетка-предшественница – клетка, находящаяся на низком уровне дифференцировки, но уже коммитированная к развитию в клетки определенной линии.
Аксиомой современной теории онкогенеза является положение, что клеткой-предшественницей стволовой злокачественной клетки является нормальная пролиферирующая соматическая клетка. Однако какая соматическая клетка явилась клеткой-предшественницей для злокачественной клетки, данной конкретной солидной опухоли – не известно.
Достоверно доказаны очень важные и никем неоспоримые утверждения:
– злокачественные клетки имеют больше сходства между собой, чем нормальные клетки между собой;
– злокачественные клетки имеют меньше различий между собой, чем различия между злокачественными клетками и нормальными клетками;
– нормальные клетки имеют меньше различий между собой, чем различия между нормальными клетками и злокачественными клетками;
– основные принципы «зарождения» стволовой злокачественной клетки, роста злокачественного очага и развития злокачественного процесса различных органов и тканей совершенно идентичны.
На этом основании можно говорить о злокачественных клетках, как об отдельной группе клеток, имеющих общее происхождение, а в совокупности со стромой, даже как об отдельной ткани в организме-носителе. В таком случае должна быть конкретная клетка, претендующая на роль «общего начала» или клетки-предшественницы первичной стволовой злокачественной клетки солидных опухолей.
При анализе всех клеток организма человека необходимо выбрать, прежде всего, те клетки, которые имеют следующие основные свойства:
1. Являются соматическими пролиферирующими клетками с продолжительным жизненным циклом (месяцы, годы).
2. Обладают автономностью: умеют свободно перемещаться по всему организму-носителю, проникать и мигрировать в органах и тканях.
3. Способны влиять на различные жизненно важные процессы: гемопоэз, гомеостаз, иммунитет, пролиферацию, созревание и дифференцировку клеток и др.
Клетками, обладающими вышеуказанными свойствами, в организме человека являются только клетки крови, из них:
– эритроциты, тромбоциты и лейкоциты – это тупиковый вариант с коротким сроком жизни (эритроциты 100–120 суток, тромбоциты около 7–10 суток, нейтрофилы менее 6–8 часов), к тому же, имеют специфические черты и достаточно ограниченные функции, поэтому не могут претендовать на роль «общего начала»;
– лимфоциты – относятся к Мононуклеарной фракции системы крови, имеют тропность к лимфоидной ткани и, как известно, унипотентные и полипотентные стволовые клетки лимфоцитопоэза являются клетками-предшественниками стволовых злокачественных клеток гемобластозов. Зрелые лимфоциты при воздействии на них специфических антигенов вновь способны трансформироваться в бластные клетки. Можно сказать однозначно, что лимфоциты прямо или косвенно участвуют в «зарождении» первичной стволовой злокачественной клетки, а также росте и развитии злокачественного процесса;
– моноциты – относятся к Мононуклеарной фракции системы крови – прослеживают свое начало от полипотентной клетки-предшественницы родоначальнице миелопоэза с последующим развитием в Моноцитарный росток (П класс), который включает в себя достаточно большое количество клеток различной потентности (полипотентные, унипотентные) и местоположения (костный мозг, сосудистое русло, ткани). Поэтому все клетки, относящиеся к Моноцитарному ростку удобнее называть Мононуклеарная фракция или Мононуклеары. С учетом особенностей Мононуклеар является наиболее вероятным кандидатом на роль «общего начала» или клетки-предшественницы первичной стволовой злокачественной клетки солидных опухолей.
Характеристика и возможности Мононуклеаров (Моноцитарный росток):
1. Морфологически недифференцируемые и дифференцируемые Мононуклеары подразделяются на три основные группы:
– костномозговые: полипотентная клетка-предшественница родоначальница миелопоэза с последующим развитием в Моноцитарный росток, унипотентная клетка-предшественница родоначальница Моноцитов, монобласт, промоноцит, моноцит;
– периферической крови: промоноцит, моноцит;
– тканевые: промоноцит, моноцит, макрофагальный бласт, промакрофаг, макрофаг.
Промоноцит и Моноцит присутствуют во всех трех группах клеток и являются промежуточным вариантом развития от костномозговой полипотентной клетки-предшественницы родоначальнице миелопоэза с последующим развитием в Моноцитарный росток (П класс) до органо- и тканеспецифического Макрофага, как конечного варианта развития.
2. Кроветворение в красном костном мозге, это единственный функционирующий очаг интенсивной пролиферации, который сохранился с эмбрионального периода развития и функционирует у взрослого человека.
3. Мононуклеары являются представителями клеток иммунокомпетентной системы и одновременно играют решающую роль в регуляции нормального гемопоэза. Мононуклеары могут ингибировать гемопоэз с помощью межклеточных взаимодействий и посредством выделения различных иммунных и не иммунных гуморальных факторов.
4. Образование клеток Моноцитарного ростка может происходить на любом этапе дифференциации от полипотентной стволовой кроветворной клетки до промиелоцита. Отличаются ли друг от друга Моноциты и Макрофаги, образовавшиеся из различных субпопуляций и каковы их специфические функции, пока не ясно.
5. Костномозговые Мононуклеары способны выходить из костного мозга в периферическую кровь, циркулировать в периферической крови по всему организму, проникать из кровеносного русла в любые органы и ткани и мигрировать в них – перемещаться в межклеточном пространстве.
6. Мононуклеар периферической крови в нормальных условиях созревает, перед тем как проникнуть в ткани, но при воспалении сроки пребывания его в периферической крови значительно сокращены, поэтому в ткани проникают его не зрелые формы, способные к активной пролиферации.
7. Тканевые Мононуклеары, это единственные клетки в организме человека, которые в нормальных условиях могут трансформироваться в другую бластную клетку – макрофаальный бласт с последующей дифференцировкой в Макрофаг.
8. Мононуклеар периферической крови, попадая в ткани, не обязательно трансформируется в Макрофаг, он может превратиться и в клетки микроокружения, например, в эпителиоидную клетку (мезенхимально-эпителиальный переход).
9. Будучи гистогенетически единой, кроветворная система в своем функционировании характеризуется определенной независимостью поведения отдельных ростков кроветворения, поэтому изначально Мононуклеары характеризуются независимостью поведения – автономностью.
10. Мононуклеары сохраняют способность к делению на всех этапах своего развития и имеют возможность трансформироваться в первичную стволовую злокачественную клетку.
11. Злокачественные клетки, подобно Мононуклеарам, обладают многими активными свойствами: влияют на пролиферацию, дифференцировку и функциональную активность различных клеток; выработку факторов роста; размножение в геле без подложки; сниженную адгезию; пониженное контактное торможение; влияние на гемопоэз; влияние на свертывающую систему крови; влияние на клеточный и гуморальный иммунитет и др.
Таким образом, тканевые Мононуклеары (Промоноцит и Моноцит), вполне могут претендовать на роль «общего начала» или клетки-предшественницы первичной стволовой злокачественной клетки солидных опухолей.
4. ГЕНОТИПИЧЕСКИЕ ИЗМЕНЕНИЯ КОСТНОМОЗГОВОГО МОНОНУКЛЕАРА
Мононуклеар, как и любая соматическая клетка, состоит из трех основных компонентов: клеточной мембраны, цитоплазмы и ядра (Рис. 9).
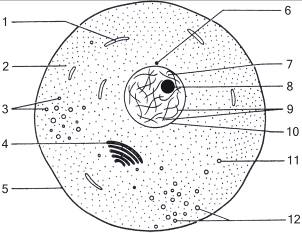
Рис. 9. Обобщенная схема животной клетки (Н. Грин, У. Стаут, Д. Тейлор, 1990, Т.1, стр. 211): 1 – митохондрии, 2 – цитоплазма, 3 – питательные гранулы, 4 – аппарат Гольджи, 5 – клеточная мембрана, 6 – центриоль, 7 – кариоплазма, 8 – ядрышко, 9 – хроматин, 10 – ядерная мембрана, 11 – лизосомы, 12 – секреторные гранулы.
Характеристика ядра клетки
Ядро – наиболее важная структура клетки, в нем сосредоточена основная масса ДНК, являющаяся носителем генетической информации. Кроме того, ядро регулирует всю «повседневную» жизнедеятельность клетки.
Содержимое ядра:
1. Хромосомы, в которые входит фактически вся ДНК ядра, видны как дискретные тела, когда клетка находится на стадии активного деления. В состоянии покоя в периоды между делениями клетки (интерфаза) хромосомы могут быть не видны.
2. Ядрышко – особое тельце, в котором сосредоточена большая часть РНК в покоящемся или интерфазном ядре.
3. Ядерная плазма (кариоплазма) – это жидкость, содержащая соли и белки, но не содержащая нуклеиновых кислот.
Ядро ограничено ядерной мембраной (кариолеммой), состоящей из двух липопротеидных слоев. Наружная мембрана связана с рибосомами, а к внутренней – тесно прилежит хроматин кариоплазмы. Наружная и внутренняя мембраны сливаются в области ядерных пор, через которые осуществляется транспорт белков и РНК. Поры ядерной мембраны заполнены белковым конгломератом, который изолирует кариоплазму от цитоплазмы, поэтому состав кариоплазмы, в том числе по содержанию ионов, отличается от состава цитоплазмы.
Ген – это участок ДНК, содержащий программу построения только одного определенного белка по формуле «Один ген – один белок». Информация, содержащаяся в гене, передается в цитоплазму посредством матричной, или информационной РНК. Если контакт ядра с цитоплазмой прекращается, то скорость всех реакций в клетке постепенно замедляется, и она погибает. В период деления происходит «ремонт», воспроизведение и удвоение молекул ДНК, что позволяет передать дочерним клеткам одинаковый в количественном и качественном отношении объем генетической информации.
Инициация костномозгового Мононуклеара
Miller E., Miller J. (1966): предложили модель онкогенеза, в последствие названную, как «теория молекулярно-генетических механизмов многостадийного канцерогенеза». Процесс происходит в две стадии: инициации и промоции.
Мы считаем, что предложенная «теория молекулярно-генетических механизмов многостадийного канцерогенеза» в действительности является только частью периода «зарождения» первичной стволовой злокачественной клетки. Тогда как модель онкогенеза включает в себя более широкое толкование, о чем будет изложено ниже.
1-я стадия (инициации от лат. initium – начало) периода «зарождения» стволовой злокачественной клетки: под канцерогенным воздействием (ионизирующее излучение, эндо- и экзоканцерогены, вирусы) наряду с относительно нейтральными повреждениями генома, могут происходить значимые мутации в онкогенах и антионкогенах. При этом возникает характерный спектр нарушений на генном, хромосомном и геномном уровнях: амплификации (увеличение копийности генов), делеции, инсерции, транслокации, микромутации (точковые замены, микроделеции, микроинсерции) и др.
Важными положениями 1-ой стадии (инициации) являются:
1. Воздействие инициатора первично.
2. Доза инициатора влияет на частоту «зарождения» злокачественной клетки.
3. Инициация необратима.
4. Инициатор может быть применен однократно и кратковременно.
5. Инициатор воздействует самостоятельно.
Известно, что генотипические изменения ядерной ДНК наиболее часто возникают в процессе митоза в зоне активной пролиферации. В организме человека наиболее интенсивная пролиферация происходит в красном костном мозге, и здесь сохранился наиболее недифференцированный состав стволовых клеток, приближенных к эмбриональным.
Интенсивность пролиферации в красном костном мозге может значительно возрастать при хроническом воспалении. Это увеличение отмечается в костном мозге, периферической крови, селезенке, лимфатических узлах, перитонеальных полостях. При ускоренной пролиферации образование клеток Моноцитарной серии может происходить на любом этапе дифференциации от полипотентной стволовой кроветворной клетки до Промиелоцита. Отличаются ли друг от друга Моноциты и Макрофаги, образовавшиеся из различных субпопуляций клеток-предшественников и каковы их специфические функции – не известно.
Генотипические изменения ядерной ДНК
Boveri T. (1914): в основе малигнизации лежит мутация, возникающая под влиянием эндогенных и экзогенных канцерогенных веществ и облучения.
Во время усиленной пролиферации при стимулированном гемопоэзе, под канцерогенным воздействием (ионизирующее излучение, эндо- и экзоканцерогены, вирусы), в клетке во время митоза может проявиться свойство «геномной нестабильности».
Наряду с относительно нейтральными повреждениями генома, могут происходить значимые мутации в онкогенах и антионкогенах. При этом возникает инициированная клетка, имеющая характерный спектр нарушений на генном, хромосомном и геномном уровнях: амплификации (увеличение копийности генов), делеции, инсерции, транслокации, микромутации (точковые замены, микроделеции, микроинсерции) и др.
Генотипические изменения могут возникнуть на различной стадии гемопоэза и чем выше согласно схеме кроветворения, тем большей потентностью будет обладать стволовая злокачественная клетка. Поэтому важно знать не только какие возникли генотипические изменения ядерной ДНК, но и на каком уровне гемопоэза они возникли, т.е. важен не только характер генотипических изменений (спектр нарушения) ядерной ДНК, но и уровень генотипических изменений (класс по схеме кроветворения).
Как было сказано выше, все стадии гемопоэза подразделяют на шесть основных классов. В каждом конкретном случае при канцерогенном воздействии возникают различные генотипические изменения ядерной ДНК, но только на одном из вышеуказанных классов гемопоэза. Для «зарождения» стволовой злокачественной клетки солидных опухолей наиболее вероятными являются следующие уровни потентности:
1. II класс – полипотентная клетка-предшественница родоначальница миелопоэза, общая для 4 ростков (КОЕ-ГЭММег) с последующим развитием в направлении Моноцитарного ростка.
2. III класс – унипотентная клетка-предшественница родоначальница Моноцитов (КОЕ-М).
Необходимо подчеркнуть, что генотипические изменения ядерной ДНК, происходят по рецессивному признаку, поэтому в красном костном мозге эти изменения никоим образом себя не проявляют. В процессе дифференцировки, генотипические изменения закладываются в генетическом аппарате будущего Промоноцита и Моноцита, которые морфологически не отличается от нормального костномозгового Мононуклеара.
Существует еще генетический или наследственный путь передачи генотипических изменений ДНК ядра клетки, когда «кровным» родственникам в 1–15% случаев передается предрасположенность к развитию злокачественного заболевания. В этом случае генотипические изменения ядерной ДНК костномозгового Мононуклеара уже имеются и канцерогенное воздействие не обязательно или является пусковым механизмом для их реализации.
Приобретение генотипических изменений ядерной ДНК для Мононуклеара не является решающим в возможности трансформации в стволовую злокачественную клетку. Генотипически измененный Мононуклеар может делиться (митоз), созревать, дифференцироваться, преобразовываться в клетки микроокружения, трансформироваться в Макрофаг. И хотя необратимые генотипические изменения Мононуклеара являются абсолютной необходимостью, для трансформации в стволовую злокачественную клетку этого совершенно недостаточно.
Генотипические изменения ДНК ядра Мононуклеара будут терпеливо ждать до тех пор, пока не возникнут оптимальные условия для их проявления и тогда «зародившиеся» потомки – стволовые злокачественные клетки, будут властвовать в организме-носителе. Но для «зарождения» стволовой злокачественной клетки, еще необходимы эпигенетические изменения, которые могут происходить, только если клетка окажется в определенном микроокружении, характеризующимся, как «суперусловия».
Таким образом, в красном костном мозге при канцерогенном воздействии во время ускоренного гемопоэза может возникнуть инициированная клетка, имеющая различные генотипические изменения ядерной ДНК.