 | |
МЕНЮ
- Главная
- Языкознание филология
- Финансовые науки
- Управленческие науки
- Товароведение
- Технология
- Теплотехника
- Теория организации
- Теория государства и права
- Таможенная система
- Схемотехника
- Строительство
- Страхование
- Статистика
- Религия и мифология
- Психология и педагогика
- Промышленность производство
- Медицинские науки
- Медицина
- Краеведение и этнография
- Компьютерные науки
- История
- Искусство и культура
- Информатика
- Инвестиции
- Издательское дело и полиграфия
- Зоология
- Журналистика
- Естествознание
- Деньги и кредит
- Делопроизводство
- Гражданское право и процесс
- Государство и право
- Геополитика
- Геология
- Геодезия
- География
- Военная кафедра
- Ветеринария
- Валютные отношения
- Бухгалтерский учет и аудит
- Ботаника и сельское хоз-во
- Биржевое дело
- Биология и химия
- Биология
- Безопасность жизнедеятельности
- Банковское дело
- Астрономия
- Астрология
- Архитектура
- Арбитражный процесс
- Административное право
- Авиация и космонавтика
- Карта сайта
Дипломная работа: Параметры функционирования митоКАТФ у животных с различной устойчивостью к гипоксии, а также у крыс, адаптированных к кислородному голоданию
Причем, необходимо заметить, что большая часть перекрывающейся последовательности приходится на Р-домен кальрегулина, способного связывать нуклеотиды. Также, в перекрывающуюся последовательность включены высоконсервативные остатки триптофана, характерные для кальрегулинов в данных сайтах первичной последовательности, и стерически идентично расположенные в первичной аминокислотной последовательности кальрегулина консервативные антипараллельные бета-листы, формирующие Р-домен.
Следует особенно отметить тот факт, что в составе прекурсорного белка и кальретикулинов различных типов имеется характерная гидрофобная сигнальная последовательность из 17 аминокислотных остатков, MLLSVPLLLGLLGLAAA (1-17), посттрансляционно удаляемая протеолитическим расщеплением [Murthy et al., 1990] (Рис.16). Эта последовательность отсутствует в изучаемом нами белке с м.м. 55 кДа. Интересно также то, что N-концевая последовательность с 17 по 27 аминокислотный остаток DPAIYFKE кальретикулина совпадает с последовательностью N-концевого участка 55 кДа белка, структура которого была определена ранее в нашей лаборатории реакцией химической деградации по Эдману [неопубликованные данные] (Рис.16).
Кроме того, С-конец прекурсорного белка, как и всех кальретикулинов, содержит сигнальную последовательность KDEL, определяющую локализацию этих белков в ретикулюме [Schweizer et al., 1993], в то время как у изучаемого 55 кДа белка такой последовательности нет (Рис.15, 16), что, вероятно, свидетельствует о том, что белок с м.м. 55 кДа локализуется не в ретикулюме.
1 MLLSVPLLLG LLGLAAADPA IYFKEQFLDG DAWTNRWVES KHKSDFGKFV
51 LSSGKFYGDQ EKDKGLQTSQ DARFYALSAR FEPFSNKGQT LVVQFTVKHE
101 QNIDCGGGYV KLFPGGLDQK DMHGDSEYNI MFGPDICGPG TKKVHVIFNY
151 KGKNVLINKD IRCKDDEFTH LYTLIVRPDN TYEVKIDNSQ VESGSLEDDW
201 DFLPPKKIKD PDAAKPEDWD ERAKIDDPTD SKPEDWDKPE HIPDPDAKKP
251 EDWDEEMDGE WEPPVIQNPE YKGEWKPRQI DNPDYKGTWI HPEIDNPEYS
301 PDANIYAYDS FAVLGLDLWQ VKSGTIFDNF LITNDEAYAE EFGNETWGVT
351 KAAEKQMKDK QDEEQRLKEE EEDKKRKEEE EAEDKEDEDD RDEDEDEEDE
401 KEEDEEDATG QAKDEL
Рисунок 14. Последовательность прекурсорного белка. Сигнальная последовательность, характерная для кальретикулинов, выделена серым цветом, N-концевая последовательность, гомологичная таковой у исследуемого белка-канала с м.м. 55 кДа – жирным шрифтом, сигнальная последовательность KDEL, остутствующая в изучаемом белке – серым цветом и жирным шрифтом
Следует также отметить, что кальретикулины содержат ряд высоко консервативных последовательностей [Fliegel et al., 1989]. Согласно результатам MS-MALDI-TOF/TOF анализа, ряд гомологичных с белком предшественником участков цепи белка с м.м. 55 кДа являются высококонсервативными для белков семейства кальретикулинов.
Возможно, исследуемый нами белок, гомологичный кальрегулину, проходит альтернативные стадии пострансляционных модификаций и созревания при биосинтезе. Это, вероятно, позволяет ему встраиваться во внутреннюю мембрану митохондрий беспрепятственно и без значительных потерь энтропии и внутренней энергии при смене гидрофильной среды на гидрофобную. Не исключено также дополнительное стерическое и физико-химическое влияние на химические свойства и конформационную норму со стороны компонентов мембраны митохондрий. Учитывая все вышесказанное, на данном этапе исследования структуры митоКАТФ, можно утверждать лишь, что белок с м.м. 55 кДа является белком, обладающим высокой степенью структурной и функциональной гомологии с кальрегулином.
Учитывая все вышесказанное, на данном этапе исследования структуры митоКАТФ, можно утверждать лишь, что белок с м.м. 55 кДа является белком, обладающим высокой степенью структурной и функциональной гомологии с кальрегулином.
5.3 Ингибиторный анализ активности митоКАТФ канала с использованием антител, полученных на белок с м.м. 55 кДа
В нашей лаборатории были получены косвенные доказательства гетеромультимерного строения этого канала, а также данные о том, что белок с м.м. 55 кДа является канальной субъединицей митоКАТФ [Григорьев, 1999; Негода, 2004; Mironova et al., 2004]. В то же время, результаты исследования гомологичности структуры белка-канала с м.м. 55 кДа, выделенного из МХ печени крысы, показали высокий процент гомологии исследуемого белка с кальрегулином. Таким образом, возникла необходимость доказательства принадлежности белка с м.м. 55 кДа к системе митохондриального АТФ-зависимого транспорта калия.
Для прямого доказательства принадлежности каналообразующего белка с м.м 55 кДа к митохондриальной системе АТФ-ингибируемого транспорта К+, в работе на этот белок были получены поликлональные антитела (АТ) и проведен анализ их влияния на АТФ-зависимый транспорт калия в интактных МХ.
Предварительные данные по влиянию АТ к 55 кДа белку на транспорт К+ в МХ были получены в лаборатории ранее [Скарга и др., 1986]. Однако в о время, белок не был идентифицирован как АТФ-зависимый К+ канал и методы его очистки были несовершенны. Кроме того, не все использовавшиеся ранее модели отражали работу АТФ-зависимого митоходнриального калиевого кнала. Не было также проведено сравненительное исследование по влиянию АТ к 55 кДа белку на другие функции МХ.
5.3.1 Определение степени чистоты белка, используемого для иммунизации
Для получения поликлональных специфических антител на белок-канал необходимо было получить гомогенный белок с м.м. 55 кДа. Этот белок выделяли из МХ печени крысы методом водно-этанольной экстракции [Миронова и др., 1981; 1996(I)]. Дальнейшую очистку белка проводили методом ионообменной хроматографии с использованием ДЭАЭ-целлюлозы (п.п. 2 «Материалов и методов»). Каналообразующий белок элюировался с колонки 250 мМ KCl. Чистота данной фракции определялась ДДС-ПААГ электрофорезом [Laemmli, 1979] (Рис.5). Для обнаружения активной фракции, все фракции, элюированные с колонки ДЭАЭ-целлюлозы тестировали при встраивании в БЛМ (см. «Материалы и методы»). Далее активную фракцию (250 мМ KCl) диализовали против 5 мМ Трис буфера (pH 7.2) с добавлением 0.1% b-меркаптоэтанола в течение ночи при 4°С. Диализат повторно наносили на колонку ДЭАЭ-целлюлозы, аналогичную использовавшейся в первом случае. Фракции элюировали ступенчатым градиентом KCl. Все фракции тестировались на активность при встраивании в БЛМ. АТФ-ингибируемые К+-селективные каналы формировались, как правило, при встраивании белка, элюированного 250 мМ KCl.
Дополнительная очистка проводилась методом препаративного нативного электрофореза (п.п. 5.1.1. «Материалов и методов») в 10%-ом ПААГ. На конечной стадии очистки, исследуемый белок элюировали из геля, концентрировали методом обратного диализа на ПЭГ и проверяли на ДДС-ПААГ электрофорезе (Рис.17). Электрофоретическая подвижность белка соответствовала массе ~55 кДа.
|
|
|
|
|
|

Рис. 15. Результат ДДС-ПААГ электрофореза фракции, полученной после нативного электрофореза и содержащей белок с м.м. 55 кДа. 1 – стандарт молекулярных масс, 2 – фракция митоКАТФ канала после нативного электрофореза
Полученным электрофоретически гомогенным белком с м.м. 55 кДа модифицировали искусственные бислойные мембраны. Было показано, что данный белок обладает селективной К+-транспортирующей активностью и ингибируется физиологическими концентрациями АТФ (Рис.18).
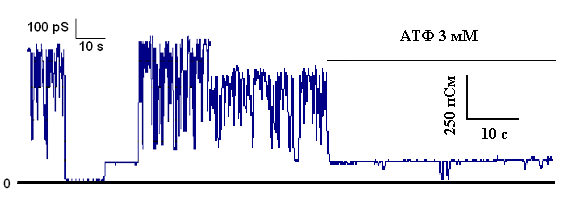
Рисунок 16. К+-транспортирующая АТФ-ингибируемая активность белка с м.м. 55 кДа, реконструированного в искусственную мембрану. Среда инкубации содержала: 20 мМ Трис, 100 мМ KCl, рН 7.2
Электрофоретически гомогенный АТФ-ингибируемый К+-транспортирующий белок с м.м. 55 кДа и замораживали и накапливали для иммунизации. При выделении вышеописанным методом из 100 г печени крыс получали 30-70 мкг очищенного белка. Иммунизацию кроликов проводили при накоплении 0.2-0.4 мг белка.
5.4 Иммунизация и определение титра полученных антител
Перед началом иммунизации отбирали 30-40 мл крови каждого животного для приготовления нормальной контрольной сыворотки. Для получения антител к АТФ-ингибируемому К+-транспортирующему белку с м.м. 55 кДа кроликов иммунизировали электрофоретически чистым белком. С целью устранения индивидуальных различий в получаемых анителах иммунизировали двух кроликов массой 2.6-2.8 кг.
Схема эксперимента была подобрана в соответствии с требованиями Декларации совета Европейского Союза 86/609/EEC и представлена в таблице 8. Инъекции делали подкожно в область лопаток в 3 подхода. Интервал между инъекциями 10-15 дней.
В ходе иммунизации кроликов по представленной выше схеме были получены антитела к белку с м.м. 55 кДа, выделенному из МХ печени крысы. Определение титра антител, полученных на белок-канал с м.м. 55 кДа проводили методом непрямого дот-анализа (см. «Материалы и методы»). Средние значения титра антител указаны в таблице 8.
Таблица 2. Схема иммунизации кроликов АТФ-зависимым К+-транспортирующим белком м.м. 55 кДа.
| Номер животного | Кролик №1 | Кролик №2 | ||||
| Номер иммунизации | 1 | 2 | 3 | 1 | 2 | 3 |
| Количество белка, мкг | 114 | 70 | 30 | 100 | 114 | 38 |
| Суммарное количество белка, мкг | 214 | 252 | ||||
| Титр после трех иммунизаций | 1:3200 | 1:1600 | ||||
5.4.1 Определение специфичности полученных антител
Специфичность антител на белок с м.м. 55 кДа определяли методом Вестерн-блот. Для этого выделенный из МХ печени крысы и очищенный АТФ-чувствительный К+-транспортирующий белок с м.м. 55 кДа наносили на ДДС-ПААГ электрофорез с 10%-ым ПААГ гелем с нагрузкой на полосу 10 мкг. На другую полосу наносили 10 мкг тотальной белковой фракции МХ. Порядок нанесения образцов дублировался на том же геле. После электрофоретического разделения белков половина геля, содержащая оба образца и стандарт молекулярных масс, окрашивалась серебрением [Shevchenko et al., 1996]. Вторая половина использовалась для переноса на нитроцеллюлозную мембрану с последующей обработкой полученными антителами (п.п. 5.2.2. «Материалов и методов»). При этом исследуемую антисыворотку разводили 1:300, вторичные антитела – 1:500.
|
|
|
|


Рисунок 17. А. ДСН-электрофорез фракций
1 – очищенный АТФ-чувствительный К+-транспортирующий белок с м.м. 55 кДа, 2 – молекулярные стандарты массы белков, 3 – суммарный белок МХ. Б. Вестерн-блот анализ: 1 – очищенный АТФ-чувствительный К+-транспортирующий белок с м.м. 55 кДа, 2 – суммарный белок МХ.
Показано, что полученные в работе антитела специфически связываются только с белком с м.м. 55 кДа, и не вязываются ни с одним из массы белков, находящихся в МХ (Рис.19). Таким образом, в работе были получены специфические поликлональные антитела на белок с м.м. 55 кДа.
Этим же методом было установлено, что поликлональные антитела на исследуемый белок-канал, полученный из МХ печени крысы, не взаимодействует с той же концентрацией аналогичного белка с м.м. 55 кДа, выделенного тем же методом из МХ сердца крысы. Данный факт указывает на то, что исследуемый белок с м.м. 55 кДа тканеспецифичен.
5.4.2 Выделение иммуноглобулинов G (IgG) из антисыворотки и проведение ингибиторного анализа
Полученные антитела к АТФ-чувствительному К+-транспортирующему белку с м.м. 55 кДа, выделенному из МХ печени крысы, использовали в качестве ингибитора АТФ-зависимого транспорта ионов калия в МХ печени крысы. Для этого, полученные очищенные и концентрированные антитела (иммуноглобулины G (IgG)) кроликов (см. «Материалы и методы»). Контролем служили очищенные IgG сыворотки крови Интактных (неиммунизированных) кроликов. Кроме того, в качестве контроля использовались антитела к белку с м.м. 55 кДа, подвергнутые кипячению в течение 5 минут. Ранее было показано, что такая процедура ведет к потере белком активности. IgG контрольных животных получали тем же способом, что и IgG рабочей сыворотки. В ходе выделения и очистки IgG титр антител и сродство к белку-каналу с м.м. 55 кДа существенно не изменяется. Степень чистоты выделенных из сыворотки IgG определяли методом ДДС-ПААГ электрофореза [Laemmli, 1979] (см. «Материалы и методы»).
Очищенные фракции IgG, выделенные из иммунизированных и интактных животных, использовали для ингибирования АТФ-зависимого выхода ионов К+ из МХ в присутствии ДНФ, отражающего работу митоКАТФ, и энергозависимого входа ионов К+ в МХ. Все эксперименты проводились при термостатировании (26°С) и постоянном перемешивании. После 1.5-2 минут преинкубации антител с митохондриями транспорт ионов К+ индуцировали ДНФ (при исследовании выхода К+ из МХ К+-селективным электродом) или субстратом дыхания (при определении энергозависимого входа К+ в МХ). Для повышения эффективности взаимодействия IgG с внутренней мембраной МХ создавались гипотонические условия. На рисунке 6 представлена концентрационная зависимость степени ингибирования выхода ионов К+ из МХ печени крысы в присутствии разобщителя (ДНФ). Ингибирующий эффект антител наблюдается только в случае добавления в среду инкубации МХ очищенных интактных IgG к АТФ-зависимому К+-транспортирующему белку с м.м. 55 кДа, выделенному из печени крысы. Степень ингибирования зависела от концентрации IgG и времени преинкубации.
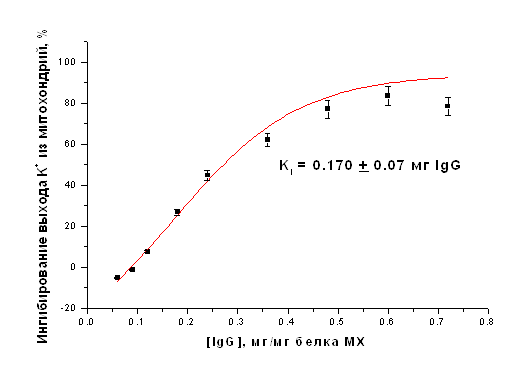
Рисунок 18. Ингибирование АТФ-зависимого ДНФ-индуцированного выхода К+ из МХ печени крысы антителами на АТФ-чувствительный К+ белок-канал с м.м. 55 кДа. Кi = 0.170+0.07 мг IgG/мг белка МХ
Максимальное ингибирование, наблюдавшееся в данных экспериментах составляло 83%. Константа полумаксимального ингибирования (Кi) составила 0.170+0.07 мг IgG/мг белка МХ (Рис. 20).
Антитела к белку с м.м. 55 кДа, выделенному из МХ печени крысы, ингибируют также энергозависимый вход ионов К+ (Рис. 21).
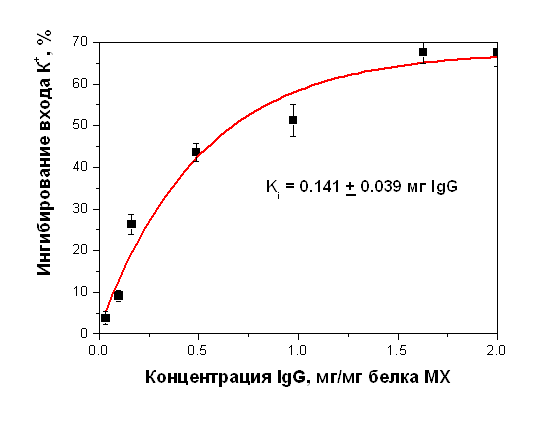
Рисунок 19. Ингибирование энергозависимого входа ионов К+ в МХ печени крысы антителами на белок с м.м. 55 кДа
Следует отметить, что IgG контрольной сыворотки и IgG, инактивированные кипячением, не влияли ни на ДНФ-индуцированный выход калия из МХ, ни на энергозависимый вход ионов (Таблица 9). Кроме того, антитела, полученные на АТФ-зависимый К+-транспортирующий белок с м.м. 55 кДа, выделенный из МХ печени крысы, не блокировали АТФ-чувствительный К+ транспорт в МХ сердца крысы, при использовании обоих методов исследования (Таблица 9).
ИНТЕРЕСНОЕ
© 2009 Все права защищены. |