 | |
МЕНЮ
- Главная
- Языкознание филология
- Финансовые науки
- Управленческие науки
- Товароведение
- Технология
- Теплотехника
- Теория организации
- Теория государства и права
- Таможенная система
- Схемотехника
- Строительство
- Страхование
- Статистика
- Религия и мифология
- Психология и педагогика
- Промышленность производство
- Медицинские науки
- Медицина
- Краеведение и этнография
- Компьютерные науки
- История
- Искусство и культура
- Информатика
- Инвестиции
- Издательское дело и полиграфия
- Зоология
- Журналистика
- Естествознание
- Деньги и кредит
- Делопроизводство
- Гражданское право и процесс
- Государство и право
- Геополитика
- Геология
- Геодезия
- География
- Военная кафедра
- Ветеринария
- Валютные отношения
- Бухгалтерский учет и аудит
- Ботаника и сельское хоз-во
- Биржевое дело
- Биология и химия
- Биология
- Безопасность жизнедеятельности
- Банковское дело
- Астрономия
- Астрология
- Архитектура
- Арбитражный процесс
- Административное право
- Авиация и космонавтика
- Карта сайта
Дипломная работа: Параметры функционирования митоКАТФ у животных с различной устойчивостью к гипоксии, а также у крыс, адаптированных к кислородному голоданию
Дипломная работа: Параметры функционирования митоКАТФ у животных с различной устойчивостью к гипоксии, а также у крыс, адаптированных к кислородному голоданию
СодержаниеСписок сокращение
Введение
Глава 1. Обзор литературы
1.1 Системы транспорта калия в митохондриях
1.1.1 Транспорт калия в митохондрии
1.1.2 Молекулярные структуры, ответственные за транспорт калия в МХ
1.1.3 Физиологическое значение транспорта калия в МХ
1.1.4 АТФ-ингибируемые калий-транспортирующие каналы
1.2 АТФ-зависимый калиевый канал цитоплазматической мембраны
1.2.1 Структурная организация цитоплазматического АТФ-зависимого калиевого канала
1.3 АТФ-чувствительный калиевый канал внутренней мембраны МХ
1.3.1 Структурная организация митоКАТФ канала
Глава 2. Модуляторы
2.1 Модуляторы митоКАТФ канала
2.1.1 Метаболические модуляторы митоКАТФ канала
2.2 Функциональная роль митоКАТФ
2.2.1 Активация митоКАТФ в развитии устойчивости организма к гипоксии
2.2.2 Механизмы защиты сердца при гипоксии, опосредованные активацией митоКАТФ
2.2.3 Феномен прерывистой гипобарической тренировки
2.3 Выделение МХ
2.3.1 Выделение МХ печени крысы
2.3.2 Выделение МХ сердца крысы
2.3.2 Выделение и очистка митоКАТФ канала
Глава 3. Изучение энергозависимого входа К+ в МХ методом спектрофотометрии
3.1 Изучение ДНФ-индуцированного выхода ионов калия из МХ
3.2 Получение и очистка антител к белку с молекулярной массой 55 кДа.
3.2.1 Подготовка белка с м.м. 55 кДа: выделение и очистка
3.3 Иммунизация и анализ препарата антител
3.3.1 Детекция специфических антител и определение титра
3.3.2 Вестерн-Блот анализ
3.4 Исследование ДНФ-индуцированного выхода К+ из митохондрий с помощью К+-селективного электрода
3.5 Реконструкция белка в БЛМ
3.6 Иммунноэлектронная микроскопия
3.7 MS-MALDI-TOF/TOF- анализ
3.8 Очистка антител к АТФ-зависимому белку с м.м. 55 кДа
3.9 Очистка антител к АТФ-зависимому белку с м.м. 55 кДа на колонке с иммобилизованным Белком А
3.10 Ингибиторный анализ с использованием антител к белку
с м.м. 55 кДа
Глава 4. Выделение комплекса цитоплазматических мембран и микросом печени крыс
4.1 Метод отбора высоко- и низкоустойчивых животных
Глава 5. Результаты и обсуждения
5.1 Параметры функционирования митоКАТФ канала у крыс с различной резистентностью, а также у животных, адаптированных к гипоксии
5.1.1 Изучение параметров дыхания и окислительного фосфорилирования в МХ печени и сердца крыс с различной резистентностью к гипоксии
5.1.2 Изучение параметров АТФ-зависимого транспорта К+ в МХ печени сердца крыс с различной резистентностью к гипоксии
5.1.2 Изучение параметров АТФ-зависимого транспорта К+ в МХ печени сердца крыс с различной резистентностью к гипоксии
5.2 Изучение структурной организации митохондриального АТФ-зависимого калиевого канала
5.2.1 Определение гомологии белка с м.м. 55 кДа методом MS-MALDI-TOF/TOF
5.3 Ингибиторный анализ активности митоКАТФ канала с использованием антител, полученных на белок с м.м. 55 кДа
5.3.1 Определение степени чистоты белка, используемого для иммунизации
5.4 Иммунизация и определение титра полученных антител
5.4.1 Определение специфичности полученных антител
5.4.2 Выделение иммуноглобулинов G (IgG) из антисыворотки и проведение ингибиторного анализа
5.5 Электронная микроскопия МитоКАТФ канала
ЗАКЛЮЧЕНИЕ
ВЫВОДЫ
ЛИТЕРАТУРА
СПИСОК СОКРАЩЕНИЙ
МитоКАТФ – митохондриальный АТФ-чувствительный калиевый канал
ЦитоКАТФ – цитоплазматический АТФ-чувствительный калиевый канал
5-ГД – 5-гидроксидекановая кислота
Глиб – глибенкламид
АТФ – аденозин-5’-трифосфат
УТФ – уридин-5’-трифосфат
УДФ – уридин-5’-дифосфат
УМФ – уридин-5’-монофосфат
ГТФ – гуанин-5’-трифосфат
МХ – митохондрии
АТ – антитела
ФН – неорганический фосфат
АФК – активные формы кислорода
KIR – inward rectifying K+ channels (канальная субъединица митоКАТФ канала)
SUR – sulphonyl urea receptor (регуляторная субъединица митоКАТФ канала,
чувствительная к сульфонилмочевинам)
PCO’s – potassium channel openers (активаторы калиевых каналов)
NFBs – nucleotide phosphate binding domains (нуклеотидсвязывающие участки)
БЛМ – бислойные липидные мембраны
ФИА - феномен ишемической адаптации
ГТ – гипоксическая тренировка
НУ – крысы, низкоустойчивые к гипоксии
ВУ – крысы, высокоусточивые к гипоксии
Введение
Митохондриальный АТФ-ингибируемый калиевый канал (митоКАТФ), осуществляющий вход калия в МХ, был обнаружен методом пэтч-кламп во внутренней мембране МХ в 1991 г. [Inoue et al., 1991]. Однако, еще в 1981 г. в лаборатории проф. Мироновой был выделен белок с м.м. 55 кДа, обладающий свойствами данного канала. [Миронова и др., 1981]. Позднее было показано, что выделенный белок-канал ингибируется физиологическими концентрациями АТФ [Paucek et al., 1992; Миронова и др., 1996 (I)].
В настоящее время достаточно хорошо исследованы биофизические свойства митохондриального калиевого канала и его физиологическая роль [Миронова и др., 1996 (I, II); Paucek et al., 1992; Inoue et al., 1991; Garlid et al., 1997; Mironova et al., 1999; 2004]. Интерес к исследованию этого канала в последнее время возрос, поскольку было показано, что он, а именно его активация, играет ключевую роль в защите миокарда при ишемии [Grover et al., 1992; Garlid et al., 1997; Vanden Hoek, 2000]. Найден целый ряд синтетических активаторов митоКАТФ, являющихся потенциальными кардиопротекторами [Gross et al., 1992; Liu et al., 1998; Sato et al., 1998; Tsai et al., 2002]. Недавно в лаборатории проф. Мироновой был обнаружен эффективный природный метаболический активатор митоКАТФ – уридин-5’-дифосфат (УДФ) [Mironova et al., 2004; Негода А.Е., 2004]. Метаболические активаторы канала имеют ряд преимуществ, по сравнению с их синтетическими аналогами, поскольку их концентрацию в клетке можно регулировать и они не обладают отрицательными побочными эффектами. Действие УДФ, как активатора К+-канала, и, следовательно, кардиопротектора, до настоящего времени не было изучено на животных.
Помимо важной роли митоКАТФ в защите миокарда от ишемических повреждений, некоторые исследователи предполагают участие активации канала в формировании устойчивости организма к кислородному голоданию [Zhu et al., 2003]. Однако прямые доказательства участия канала в адаптации к гипоксии до настоящего времени не получены, то есть, данный феномен также требует дополнительных исследований.
К настоящему времени нами разработана рабочая модель структуры и регуляции канала, в соответствии с которой митоКАТФ канал состоит из двух субъединиц – канальной (с молекулярной массой 55 кДа) и регуляторной (с молекулярной массой 63 кДа). Однако вопрос о структуре канала и возможной гомологии с другими белками пока остается открытым. Изучение структуры митоКАТФ позволит исследовать функцию и регуляцию канала на молекулярном уровне.
В связи с этим, целью данной работы было: изучить структурную организацию митоКАТФ канала, кардиопротекторное действие УДФ при ишемии миокарда, исследовать параметры функционирования митоКАТФ у животных с различной устойчивостью к гипоксии, а также у крыс, адаптированных к кислородному голоданию.
Таким образом, в работе были поставлены следующие задачи:
1) Исследовать параметры функционирования митоКАТФ у крыс с различной устойчивостью к гипоксии, а также у животных, адаптированных к недостатку кислорода.
2) Определить гомологию структуры исследуемого белка с м.м. 55 кДа аминокислотным последовательностям известных белков.
3) Получить специфические поликлональные антитела на белок-канал с м.м. 55 кДа, формирующий при встраивании в искусственные мембраны АТФ-ингибируемые К+ каналы.
4) Провести ингибиторный анализ АТФ-чувствительного транспорта калия в нативных МХ с использованием полученных антител (АТ) с целью доказательства принадлежности белка с м.м. 55 кДа к системе АТФ-зависимого транспорта К+ в МХ.
5) Исследовать иммунолокализацию МитоКАТФ канала на срезе гепатоцитов и кардиомиоцитах при помощи электронной микроскопии
Глава 1. Обзор литературы1.2 Системы транспорта калия в митохондриях
1.1.1 Транспорт калия в митохондрии
Транспорт К+ в митохондриях (МХ) имеет большое функциональное значение, так как концентрация К+ в цитоплазме значительно превышает концентрацию других катионов, и появление любой неконтролируемой проницаемости митохондриальной мембраны для К+ может представлять угрозу осмотической целостности и функциональной интегральности МХ [Brierley et al., 1983]. МХ занимают 40% внутриклеточного пространства в сердечной клетке и до 20% - в клетке печени и по данным [Rottenberg, 1973], в свободном состоянии в матриксе МХ содержится 95 нмоль К+/мг белка МХ, в слабосвязанном состоянии – 45 нмоль и в прочносвязанном – 5 нмоль. Концентрация свободного калия в цитоплазме составляет 90-120 нмоль.
Хемиосмотическая теория Митчела включает четыре постулата, последние два из которых имеют отношение к системе транспорта катионов. Согласно этим двум постулатам, внутренняя мембрана МХ: а) непроницаема для Н+, ОН- и всех прочих ионов; б) содержит ряд белков-переносчиков, осуществляющих транспорт неорганических ионов и необходимых метаболитов. Таким образом, по Митчелу, внутренняя мембрана МХ непроницаема для катионов и анионов. Однако из-за большого электрохимического потенциала [Δφ] (~ –200 мВ с матриксной стороны) одновалентные катионы (К+ и Na+) могут диффундировать через липидный бислой. Проблема такой утечки будет наиболее актуальна для ионов К+, как основного катиона цитозоля и матрикса, хотя скорость такой диффузии будет не велика.
Что касается специфических систем транспорта, то было установлено, что в МХ существуют системы: 1) электрогенного входа калия и 2) К+/Н+-антипортер [Chavez et al., 1977; Diwan, 1981; Garlid, 1980], о существовании которого говорил Митчел [Mitchell and Moyle, 1969].
В работах с использованием радиоактивного калия (42К+) было показано, что активный транспорт К+ в МХ осуществляется электрогенно и что существует специфическая К+-транспортирующая система, функционирующая подобно ионофорам, катализирующим унипорт калия [Gamble, 1957, 1962; Judan et al., 1965; Rottenberg, 1973; Chavez et al., 1977].
В качестве системы электрогенного транспорта К+ в МХ предлагались следующие варианты: 1) подвижный переносчик или канал [Jonston and Griddle, 1978; Panov et al., 1980; Jung et al., 1982]; 2) электродиффузное движение через гидрофильные поры и 3) вход калия с участием белковых транспортных систем, которые помимо своей основной функции, могут осуществлять транспорт К+ [Brierly, 1980, 1983].
1.1.3 Молекулярные структуры, ответственные за транспорт калия в МХ
В настоящее время в литературе в качестве транспортных систем, осуществляющих электрогенный транспорт калия в МХ, рассматриваются следующие структуры: 1) АТФ/АДФ-антипортер или аденинуклеотидтранслоказа (АНТ) [Panov et al., 1980; Le Quoc et al., 1988] и 2) специфические белки, которые образуют во внутренней мембране МХ канал для электрогенного входа ионов калия [Миронова и др., 1981; Diwan et al., 1988; Paucek et a., 1992]. В первом случае предполагается, что АНТ, помимо основных функций, может работать как система электрофоретического транспорта ионов калия в МХ при действии на МХ субмикромолярных концентраций ионов кальция, и как неспецифическая пора для низкомолекулярных соединений при повреждении МХ высокими концентрациями Ca2+ [Jung and Brierley, 1981; 1982; Halestrap et al., 1986].
Что касается специфических белков, следует отметить, что в лаборатории Мироновой в 1981 г. методом водно-этанольной экстракции из МХ сердца быка был выделен и очищен белок с м.м. 55 кДа, который при реконструкции в бислойные липидные мембраны (БЛМ) образовывал К+-селективные каналы проводимости [Миронова и др., 1981], которые, как было показано позднее, ингибируются физиологической концентрацией АТФ и глибенкламидом, и относятся к семейству АТФ-чувствительных калиевых каналов [Миронова и др., 1996 (I); Mironova et al., 1999]. Структура митохондриального КАТФ канала не определена [Paucek et al., 1992].
Диван с соавторами, используя детергент Тритон Х-100, из МХ мембраны печени крыс выделили белок с м.м. 53 кДа [Diwan et al., 1988]. Реконструировав его в липосомы, авторы показали, что этот белок также обладает свойствами, характерными для К+ унипортера.
Известно, что в МХ помимо системы электрогеннного входа К+ существует система электронейтрального выхода К+ в обмен на Н+ [Garlid and Paucek, 2003]. При этом внешняя МХ мембрана не препятствует дальнейшему обмену небольшими ионами с цитоплазмой.
Система электрогенного входа калия и электронейтрального К+/Н+-обменника образуют К+ цикл МХ (Рис.1).
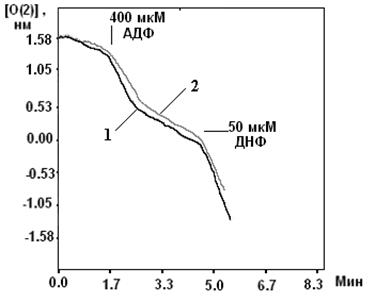
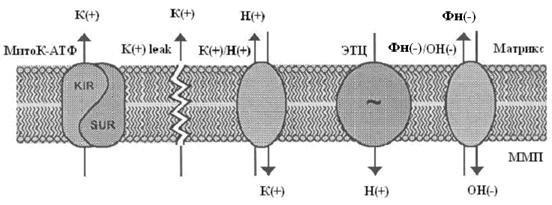
Рис. 1. МХ калиевый цикл по Garlid and Paucek, 2003. ЭТЦ – электрон-транспортная цепь; ММП – межмембранное пространство; Фн – неорганический фосфат.
Электрогенный выброс протонов электрон-транспортной системой генерирует мембранный потенциал, который, в свою очередь, стимулирует диффузию К+ в матрикс МХ («утечка калия» или К+ leak) и вход иона, опосредованный неким специфическим калиевым каналом. Такой обмен Н+ на К+ подщелачивает матрикс, вызывая вход фосфата по электронейтральному Фн-Н+ симпортеру (Фн – неорганический фосфат) (Рис.1).
Вход К+ сопровождается накоплением осмотически облигатной воды, что проявляется в набухании МХ. Излишнее их набухание может угрожать целостности этих органелл, следовательно, избыток К+ необходимо удалять. Эту функцию выполняет белок с м.м. 82 кДа - К+/Н+-антипортер [Brierley G., 1976; Garlid K., et al., 1980; 1988]. Следует отметить, что вход К+ путем диффузии слишком мал, чтобы существенно влиять на изменение объема матрикса МХ. Интенсивный вход калия в МХ обеспечивается специфическими белками, осуществляющими электрофоретический вход К+ в МХ [Mironova G., 1981; Diwan J., et al., 1988; Paucek et al., 1992].
1.1.3 Физиологическое значение транспорта калия в МХ
Предполагается, что основное физиологическое значение систем транспорта К+ в МХ связано с регуляцией объема МХ матрикса. Объем МХ определяется калиевыми токами через внутреннюю мембрану. Когда вход и выход К+ находятся в равновесии, калиевые потоки определяются электронейтральным током анионов и осмотически облигатной воды [Garlid, 1988]. Так как концентрация ионов калия в матриксе и в цитоплазме практически одинаковая, транспорт калия мало влияет на матриксную концентрацию калия, но может иметь большое влияние на объем МХ. Небольшое нескомпенсированное увеличение входа К+ в МХ может удваивать их объем в течение 1-2 минут [Garlid, 1979]. В свою очередь, увеличение матриксного объема стимулирует активность дыхательной цепи, что было показано на МХ сердца и печени [Nichols and Lindberg, 1972; Halestrap, 1989]. Окисление жирных кислот также чувствительно к изменению объема матрикса [Halestrap, 1987].
Увеличение в МХ концентрации К+ и последующее изменение их объема имеет значение и в регуляции митохондриальных процессов у зимоспящих животных [Fedotcheva et al., 1985; Бакеева и Брустовецкий, 1993], а также при адаптации животных к холоду [Nedergraard and Cannon, 1987]. В ряде работ установлено, что in vivo, гормон, участвующий в регуляции теплопродукции (тироксин), активирует системы электрогенного транспорта К+, вызывая низкоамплитудное набухание МХ [Halestrap, 1987; Shears an Brouk, 1980]. В нашей лаборатории, было показано, что активность системы электрогенного транспорта ионов калия прямо коррелирует с интенсивностью термогенеза [Федотчева и др., 1984; Миронова и др., 1986], что непосредственно связано с увеличением содержания ионов калия в МХ бурой жировой ткани и печени [Скарга, 1994]. Активацию транспорта калия при выходе животного из спячки, связывают с активацией футильного цикла К+ в МХ, приводящей к увеличению теплопродукции, до начала синтеза АТФ в МХ [Миронова и др., 1986].
Обнаружено также, что митоКАТФ канал играет существенную роль в нормальной физиологии миокарда, регулируя объем митохондрий и продукцию активных форм кислорода (АФК) [Garlid et al., 2003a].
В последнее время интерес ученых к митоКАТФ каналу вызван обнаружением его роли в защите сердца от инфаркта. Исследования были стимулированы открытием феномена кардиопротекторного действия прерывистой гипоксии, приводящей к активации митоКАТФ [Murry et al., 1986]. Установлено, что фармакологические активаторы митоКАТФ канала предохраняют сердце от ишемических повреждений [Garlid et al., 1997].
1.1.4 АТФ-ингибируемые калий-транспортирующие каналы
К+ каналы, активность которых ингибируется физиологическими концентрациями АТФ впервые были обнаружены в цитоплазматической мембране кардиомиоцитов [Noma, 1983]. С тех пор, аналогичные каналы были найдены в β клетках поджелудочной железы [Dunne and Petersen, 1991], в скелетной мускулатуре [Spruce et al., 1985; Woll et al., 1989], в нервных клетках [Jonas et al., 1990] и гладких мышцах [Standen et al., 1989]. Активность АТФ-чувствительных калиевых каналов (КАТФ каналов) связана с уровнем биоэнергетического метаболизма клетки (концентрацией АТФ) и электрическими свойствами возбудимости плазматической мембраны. Микромолярные концентрации АТФ и сульфонилмочевины – класс гипогликемических соединений, использующихся при лечении диабета, регулируют активность всех представителей семейства КАТФ каналов [Ashcroft et al., 1989]. Концентрация кальция и изменение мембранного потенциала практически не влияют на активность этих каналов [Ashcroft and Ashcroft, 1990]. В то же время, в кортикальных и гипоталамических нейронах КАТФ каналы [Ashford et al., 1989; Ashford et al., 1988] менее чувствительны к АТФ (для их ингибирования нужны миллимолярные концентрации). В эпителиальных клетках (назальные полипы) также были обнаружены КАТФ каналы с низкой чувствительностью к АТФ, но активирующиеся микромолярными концентрациями Ca2+ [Kunzelmann et al., 1989]. При этом, все вышеперечисленные каналы высокоселективны для ионов К+.
В МХ мембране также были обнаружены АТФ-зависимые К+ каналы. Так, в 1991 г. впервые с использованием метода петч-кламп [Inoue et al., 1991] на митопластах было показано, что во внутренней мембране МХ присутствуют высокоселективные по К+ каналы с невысокой проводимостью. Обнаружено, что каналы могут обратимо ингибироваться АТФ, а так же 4-аминопиридином и специфическим ингибитором цитоплазматического КАТФ канала – глибенкламидом с матриксной стороны [Inoue et al., 1991]. Однако, как было сказано выше, еще в 1981 году в лаборатории проф. Мироновой во внутренней мембране МХ был обнаружен К+-селективный АТФ-ингибируемый канал [Миронова и др., 1981; 1996; 1997; 1999], принадлежащий к семейству КАТФ каналов [Paucek et al., 1992]. Данный канал имеет те же характеристики проводимости, что и канал, обнаруженный методом пэтч-кламп. В настоящее время он интенсивно изучается, поскольку играет важную роль в функционировании клетки. Однако вопрос о его структуре требует дальнейших исследований.
1.2 АТФ-зависимый калиевый канал цитоплазматической мембраны
К настоящему времени КАТФ каналах плазматической мембраны хорошо изучены. Изначально их классификация велась по величине проводимости одиночных каналов [Ashcroft and Ashcroft, 1990], а позднее, с развитием молекулярно-биологических методик, по аминокислотному составу белковой молекулы канала [Yokoshiki et al., 1998].
1.2.1 Структурная организация цитоплазматического АТФ-зависимого калиевого канала
Молекулярно-биологические и электрофизиологические исследования последних лет позволили определить структуру цитоКАТФ канала. Исследуемый канал состоит из двух белков: KIR – inward rectifying K+ channels, формирующий пору канала, и SUR – sulphonyl urea receptor, регуляторная субъединица, придающая каналу чувствительность к модуляторам. SUR-субъединица содержит нуклеотидсвязывающие участки, локализующиеся на цитоплазматической стороне, а также участки связывающие активаторы калиевых каналов (potassium channel activators, PCOs) и MgАДФ. Каналы подсемейства KIR6.0 сами обладают чувствительностью к АТФ. Роль SUR заключается, вероятно, в облегчении доступа АТФ к АТФ-связывающему участку KIR [Tucker et al., 1997; Yokoshiki et al., 1998]. В настоящее время механизмы молекулярного взаимодействия KIR и SUR окончательно не выяснены.
Результаты исследования электрофизиологических свойств нативных КАТФ каналов позволяют предположить, что они принадлежат к классу калиевых каналов внутреннего выпрямления (KIR). В 1993 году на многие KIR каналы (KIR1.1 [Ho et al., 1993], KIR2.1 [Kubo et al., 1993], KIR3.1 [Dascal et al.,1993]) были получены клоны. Их выделили в отдельное подсемейство KIR-каналов (KIR1.0-6.0), основанное на унифицированной номенклатуре [Chandy and Gutman, 1993]. Гидрофобные участки клонированных KIR каналов содержат два мембранопересекающих сегмента М1 и М2, образующих пору. Подобно потенциалзависимым К+ каналам, KIR каналы, вероятно, формируют тетрамер [Doupnik et al., 1995; Salkoff and Jegla, 1995; Nichols et al., 1996; Nichols and Lopatin, 1997].
ИНТЕРЕСНОЕ
© 2009 Все права защищены. |