 | |
МЕНЮ
- Главная
- Языкознание филология
- Финансовые науки
- Управленческие науки
- Товароведение
- Технология
- Теплотехника
- Теория организации
- Теория государства и права
- Таможенная система
- Схемотехника
- Строительство
- Страхование
- Статистика
- Религия и мифология
- Психология и педагогика
- Промышленность производство
- Медицинские науки
- Медицина
- Краеведение и этнография
- Компьютерные науки
- История
- Искусство и культура
- Информатика
- Инвестиции
- Издательское дело и полиграфия
- Зоология
- Журналистика
- Естествознание
- Деньги и кредит
- Делопроизводство
- Гражданское право и процесс
- Государство и право
- Геополитика
- Геология
- Геодезия
- География
- Военная кафедра
- Ветеринария
- Валютные отношения
- Бухгалтерский учет и аудит
- Ботаника и сельское хоз-во
- Биржевое дело
- Биология и химия
- Биология
- Безопасность жизнедеятельности
- Банковское дело
- Астрономия
- Астрология
- Архитектура
- Арбитражный процесс
- Административное право
- Авиация и космонавтика
- Карта сайта
Реферат: Динамическое поведение мембранных систем и липидно-белковые взаимодействия
Реферат: Динамическое поведение мембранных систем и липидно-белковые взаимодействия
Динамическое поведение мембранных систем и липидно-белковые взаимодействия
Введение
Все биологические структуры по своей природе динамичны, и при рассмотрении их функций необходимо учитывать подвижность компонентов, из которых эти структуры состоят. Это относится к ферментам, полинуклеотидам и, конечно, к мембранам. В жидкостно-мозаичной модели, в центре которой находится представление о подвижности мембранных компонентов, мембрана рассматривается как некое липидное море, в котором свободно плавают глобулярные белки. За последние два десятилетия было опубликовано множество работ, посвященных количественным и качественным аспектам динамики мембранных компонентов. Во многих из них использовались спектральные методы и спектроскопия магнитного резонанса. В результате была создана весьма полезная физическая картина мембран, иллюстрирующая способы перемещения мембранных белков и липидов и их взаимодействия. Кроме того, были разработаны специальные методики, направленные на изучение динамических свойств мембран. Многое в этой области остается неясным, но уже определены основные направления будущих исследований.
Главным побудительным мотивом в изучении мембранной динамики служит ее связь с биологическими функциями мембран. Необходимым условием протекания одних ферментативных процессов является свободная диффузия мембраносвязанных компонентов в плоскости бислоя, другие же процессы могут осуществляться лишь при ограниченной подвижности мембранных компонентов. Между шероховатым и гладким эндоплазматическими ретикулумами, комплексом Гольджи и плазматической мембраной происходит быстрый обмен различными веществами, и тем не менее их состав и функции различаются. Чтобы понять суть этих и многих других биологических феноменов, необходимо прежде всего выяснить фундаментальные аспекты динамических свойств мембран. Поперечная асимметрия в распределении липидов, а возможно, и пассивная проницаемость бислоев очевидным образом связаны со скоростью трансмембранного флип-флоп-переноса липидов. Биогенез мембран зависит от скорости обмена липидов между различными мембранами. Скорость ферментативных реакций, протекающих с участием мембраносвязанных компонентов, зависит от скорости латеральной диффузии компонентов мембран. Наконец, липидно-белковые взаимодействия зависят от скорости, с которой происходит обмен липидами между ближайшим окружением белков и остальным объемом мембраны.
Диапазон движений, происходящих в мембране, весьма широк: от молекулярных колебаний с частотой порядка 1014 с до трансмембранного флип-флоп-переноса липидов, характерное время которого может достигать несколько суток. На рис. 1 в общем виде представлены некоторые из этих процессов, а также указаны временные пределы чувствительности различных биофизических методов. Величины, приведенные на рисунке, различаются на 20 порядков, поэтому термины «быстрый» и «медленный» не могут адекватно характеризовать различные типы движений. Из рисунка видно также, что одни методы позволяют получить статичную картину мембраны, поскольку характерное время соответствующих движений больше, чем время измерений, в то время как другие методы дают усредненную по времени картину, поскольку время перемещения молекул гораздо меньше, чем время измерения.
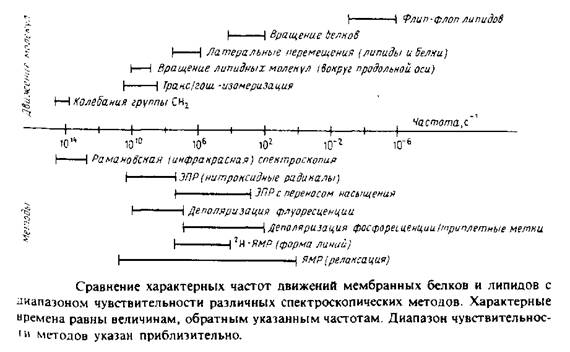
Мы рассмотрим два основных типа экспериментов. Первые основаны на использовании внутримембранных зондов для изучения текучести мембраны. Индикаторами физического состояния мембраны, а также характера липидно-белковых взаимодействий могут служить низкомолекулярные ЭПР-метки и флуоресцентные зонды.
Второй тип экспериментов направлен на прямое измерение латеральной диффузии мембранных белков или липидов и вращательной способности белков внутри бислоя. Исследовались также молекулярные взаимодействия в бислое, поскольку они влияют на динамику изучаемых молекул.
1. Некоторые простые модели движения мембранных компонентов
На рис. 2 представлены некоторые модели, использующиеся для анализа поступательного движения молекул внутри мембранного бислоя. Такие модели необходимы для интерпретации экспериментальных данных с точки зрения молекулярного движения.
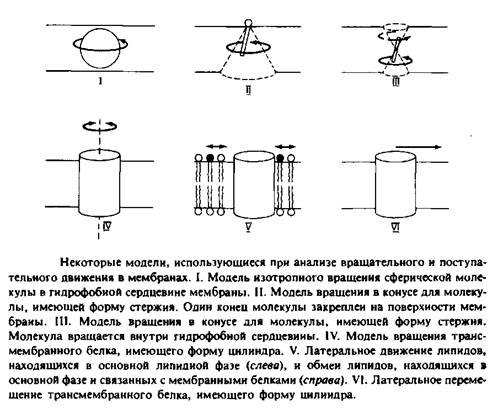
Модель изотропного вращения предполагает, что молекула вращается с одинаковой вероятностью во всех направлениях и выделенная ось вращения отсутствует. Подобное вращение будет совершать сферическая частица в непрерывной жидкой среде. Эта концепция применялась при анализе движения малых растворимых в мембране гидрофобных меток, таких, как TEMPO. Изотропная трехмерная диффузия сферической частицы рассматривается как случайное блуждание и характеризуется коэффициентом вращательной диффузии DBp. Этот параметр определяется как среднеквадратичное угловое отклонение или время релаксации.
2.Модель вращения в конусе описывает движение амфифильных меток, например производных жирных кислот. Эти молекулы можно представить в виде жестких стержней, один конец которых закреплен на поверхности мембраны. Их движение ограничивается конусом, который они описывают относительно некой оси, перпендикулярной плоскости мембраны.
3.Модель вращения в конусе используется в некоторых работах для описания движения ДФГ — гидрофобного флуоресцентного зонда, представляемого в виде жесткого стержня. В данном случае эта модель, постулирующая существование предпочтительной ориентации для ДФГ, является, безусловно, слишком упрощенной, поэтому были предложены альтернативные модели.
4. В простейшей модели вращения белков белковая молекула представляется в виде цилиндра, закрепленного в мембране и способного вращаться только вокруг оси, перпендикулярной плоскости мембраны. Такое вращение анизотропно. Время релаксации, характеризующее это движение, обычно представляется в виде 4>ц = \/. Знак «II» указывает на то, что вращение происходит
параллельно оси цилиндра.
5
и 6. Поступательное движение липидных и белковых
молекул описывается как двумерная диффузия. Особый
интерес представляет скорость обмена липидов, связанных с мембранными белками и
находящихся в основном объеме мембраны. Изотропная двумерная диффузия
характеризуется среднеквадратичным перемещением </2>,
происходящим за время ![]()
Таблица 1. Некоторые метки, использующиеся для изучения динамики мембран
Для обычной жидкости, какой является, например, вода, текучесть определяется как величина, обратная вязкости — понятному и легко измеряемому физическому параметру. Вязкость характеризует трение, возникающее между соседними слоями жидкости, которые движутся с разными скоростями. Вязкость жидкости можно оценить, измерив скорость, с которой падает мраморный шарик в жидкости. В случае мембран термин «текучесть» обычно носит скорее качественный характер: имеется в виду сопротивление, которое оказывает мембрана различным типам перемещений в ней. Как правило, для измерения текучести наблюдают за движением спиновых или флуоресцентных зондов, включенных в мембрану. Зондами обычно являются небольшие молекулы, сравнимые по размерам с мембранными фосфолипидами. Некоторые из них представлены в табл. 5.1. Теоретические основы использования спектроскопических методов для изучения молекулярных движений описаны в последующих разделах. Поскольку эти методы позволяют измерять как скорость движения, так и сопротивление этому движению, сведения о динамике и молекулярной упорядоченности даются вместе. Укажем некоторые моменты, существенные для количественной интерпретации данных по движению зондов внутри мембран.
1.Липидный бислой не является простой вязкой трехмерной гомогенной жидкостью, а представляет собой жидкую среду с низкой вязкостью, у которой состав и динамические свойства в центральной области сильно отличаются от состава и свойств периферических полярных участков.
2.Вращение зондов изотропно, как это имеет место в случае сферических частиц, не обладающих выделенной осью вращения. Часто зонды внутри мембраны имеют предпочтительную ориентацию и их движения ограничены определенными рамками. Интерпретация экспериментальных данных зависит от модели, используемой для описания молекулярного движения.
3.Локализация зондов в мембране может различаться. Например, зонд может быть связан с белковой молекулой или белковыми агрегатами, располагаться внутри липидного домена, который может находиться в различных физических состояниях.
Для оценки текучести определяют спектроскопические параметры низкомолекулярных зондов. К таким параметрам относятся: 1) время вращательной корреляции спиновых меток или флуоресцентных зондов; 2) параметр упорядоченности; 3) стационарная анизотропия флуоресцентных зондов; 4) коэффициент распределения зондов между мембранной и водной фазами. Ясно, что текучесть мембраны, оцененная только по одному параметру, не может служить достаточно полной характеристикой физического состояния мембраны. И все же измерение отдельных параметров весьма полезно, особенно для характеристики изменений физического состояния мембраны, обусловленных, например, изменениями температуры, давления, содержания холестерола, фосфолипидного или ионного состава. Обычно текучесть, измеряемая с помощью спиновых меток и флуоресцентных зондов, однозначно связана с упаковкой липидов в мембране. Воздействия, приводящие к уменьшению площади, приходящейся на одну липидную молекулу, такие, как увеличение гидростатического давления, понижение температуры или добавление холестерола к фосфолипидам в жидкокристаллическом состоянии, вызывают уменьшение текучести. Это согласуется с теорией свободного объема, согласно которой текучесть и плотность связаны между собой обратной зависимостью. Чем более плотная упаковка характерна для мембраны, тем более ограниченным будет движение зонда. Такой подход приемлем при рассмотрении текучести большинства неассоциированных жидкостей.
2. Физиологическое значение текучести мембран
Обычно биомембраны находятся в жидкокристаллическом состоянии, и, по-видимому, поддержание такого состояния очень важно для их функционирования. При переходе мембраны из жидкокристаллической фазы в фазу геля текучесть уменьшается примерно на два порядка. Структурные и динамические свойства бислоя, находящегося в фазе геля, совершенно несовместимы с организацией и правильным функционированием белковых компонентов в мембране. Впрочем, из этого правила имеются несколько исключений. Это, например, полукристаллические области пурпурных мембран Н. halobium, содержащие бактериородоп-син.
Возможно, наиболее яркое доказательство того, что текучесть, измеряемая с помощью спиновых меток или флуоресцентных зондов, играет важную физиологическую роль, получено в исследованиях по адаптации различных организмов к внешним экстремальным воздействиям. Подобные явления наблюдаются чаще всего при изучении термического стресса, когда микроорганизмы, растения, пойкилотермные или зимующие животные подвергаются воздействию низких температур. Адаптация заключается в изменении липидного состава мембран, а именно — в увеличении содержания ненасыщенных липидов или уменьшении средней длины ацильной цепи. Подобные изменения ведут к уменьшению плотности упаковки липидов в мембране и, таким образом, поддерживают текучесть мембраны. Текучесть мембран может быть критичной для одной или более мембранных функций, но каков механизм этого феномена на молекулярном уровне — неизвестно.
2.1 Характер и скорость движений, измеряемых с помощью 2Н-ЯМР, ЭПР и флуоресцентных зондов
Необходимо иметь представление о том, что именно измеряется с помощью мембранных зондов; особенно важно знать, каково отношение между временным интервалом, в котором наблюдается изучаемый тип движения, и временной шкалой, характерной для данного метода. Рассмотрим общий случай, который одинаково применим к ЭПР, измерению флуоресценции или 2Н-ЯМР. Все три спектроскопических метода чувствительны к ориентации молекул относительно лабораторной системы координат. Спектр 2Н-ЯМР чувствителен к ориентации связи С—D относительно приложенного поля. Это связано с тем, что градиент локального поля, векторно суммируясь с внешним полем, дает результирующую, которая и улавливается дейтерием. Аналогичная ситуация характерна и для ЭПР. Спектр ЭПР зависит от ориентации нитроксидной связи N— О, которую содержат большинство обычно используемых зондов, относительно приложенного магнитного поля. В случае флуоресцентной спектроскопии измеряемая поляризация испускаемого света зависит от ориентации дипольного момента перехода молекулы относительно направления, определяемого используемым поляризатором.
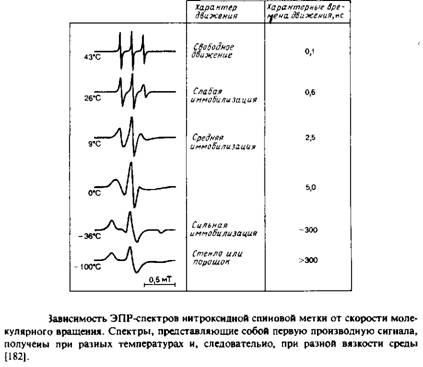
Если молекулы образца ориентированы случайным образом и полностью неподвижны, то каждая субпопуляция, характеризующаяся определенной ориентацией, будет иметь свой уникальный спектр ЭПР или ЯМР, а спектр всего образца будет равен простой сумме этих спектров с учетом их вкладов, определяемых концентрацией соответствующих молекул. Суммарный спектр называется «порошковым» или «твердостекольным» и обычно весьма широк. Такой спектр сильно отличается от спектра образца, в котором молекулы также ориентированы случайным образом, но характерное время их движений гораздо меньше времени измерения. В рассматриваемых нами примерах соответствующие частоты составляют 105 с-1 для 2Н-ЯМР и 108 с"1 для ЭПР. Каждая молекула, совершающая быстрое изотропное движение, воспринимает некое среднее магнитное поле, но при этом для всех молекул это среднее поле одинаково, поэтому пики в результирующем спектре очень острые. Примеры ЭПР-спектров приведены на рис. 5.3. Поскольку характерные времена измерения для методов ЭПР и ЯМР различаются, молекула со временем корреляции 106 с будет казаться неподвижной в первом случае и очень быстро движущейся во втором.
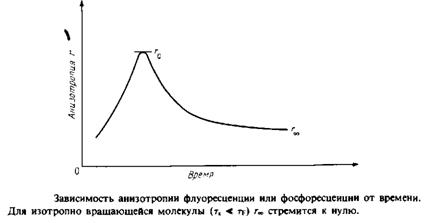
Метод измерения деполяризации флуоресценции состоит в том, что образец облучают поляризованным светом и спустя несколько наносекунд регистрируют испускаемый свет. Если молекулы остаются неподвижными в течение времени между процессами поглощения света и его испускания, то поляризация испускаемого света будет определяться в основном поляризацией света возбуждающего. Если же молекула совершает быстрое изотропное вращение, то вектор напряженности электрического поля испускаемого света не будет иметь предпочтительной ориентации, поскольку поляризация будет равна нулю. Значения поляризации, находящиеся между этими двумя крайними величинами, соответствуют или медленным движениям, при которых популяция молекул не успевает достичь полностью разупорядоченного состояния за время 10-8 с, или быстрым движениям, ограниченным в пространстве и, следовательно, не допускающим полную разупорядоченность.
Все три спектроскопических метода используются для нахождения параметра упорядоченности S:
![]()
cos20 — это усредненная по времени ориентация молекулы относительно нормали к плоскости мембраны. Время, за которое происходит усреднение, зависит от метода: 10~5 с для 2Н-ЯМР, 10~8 с для ЭПР и деполяризации флуоресценции. В случае 2Н-ЯМР этот параметр чувствителен к медленным движениям, совершающимся за времена порядка 10"5 с. Метод ЭПР или деполяризации флуоресценции регистрирует процессы, совершающиеся за время около 10"8 с, а информация о более медленных движениях утрачивается.
Данные о динамических свойствах мембран можно получить с помощью всех трех методов и более прямым способом. Так, из данных ЯМР можно найти времена релаксации, которые в свою очередь зависят от характера молекулярных движений. Из спектра ЭПР можно при определенных условиях найти время вращательной корреляции зонда. Измеряя зависимость степени деполяризации флуоресценции от времени, можно получить данные о динамическом состоянии зонда, а не только оценить степень ограниченности его движения.
2.2 Спиновые метки, использующиеся при изучении текучести мембран
ЭПР — это очень ценный метод изучения мембран. Обычно в качестве меток используются стабильные парамагнитные соединения, содержащие нитроксидный радикал. Неспаренный электрон при наложении сильного магнитного поля переходит с одного энергетического уровня на другой под действием микроволнового излучения. Метод очень чувствителен: обычно спектр регистрируется при концентрации спиновых меток около 10"*М в 50 мкл образца. Спектр ЭПР, как правило, представляют в виде первой производной от спектра поглощения.
В спектре нитроксидного радикала имеются три пика, отвечающие спин-спиновым взаимодействиям неспаренного электрона и ядра атома азота. И положение спектра, и расщепление, обусловленное спином ядра, зависят от ориентации молекулы относительно внешнего поля. Как мы уже говорили в предыдущем разделе, это означает, что характер спектра зависит от характера вращения молекулы. На рис. 5.3 приведены спектры для самых разных случаев — от свободного вращения до полной неподвижности. Для того чтобы найти тк в предположении, что вращательное движение молекулы изотропно, можно использовать простое уравнение, в которое входит высота пиков. Этот подход наиболее применим для такой метки, как TEMPO.
Спиновые метки — производные жирных кислот или фосфолипидов, — конечно, не вращаются изотропно, и в этом случае для нахождения параметра упорядоченности S из величины расщепления между линиями спектра используется вторая простая формула. Предположив, что такие метки имеют форму жесткого стержня, можно из величины параметра S оценить максимальное отклонение зонда от нормали к поверхности мембраны.
Многие предположения, которые делаются при нахождении из спектров ЭПР времен вращательной корреляции или параметра S, снижают ценность этого метода для определения количественных характеристик молекул. Эти параметры тем не менее позволяют получить качественное представление о поведении мембраны. Результаты измерения степени упорядоченности спиновых меток, фиксированных в мембране на разной глубине, свидетельствуют об увеличении неупорядоченности в направлении от поверхности мембраны к ее центральной области. Качественно такая же картина получена с помощью 2Н-ЯМР. Во многих исследованиях проводилось сравнение текучести мембран, которую определяли по данным о величинах тк и S для спиновых меток, при различных возмущающих воздействиях.
2.3 Флуоресцентные зонды, используемые для измерения текучести мембран
Для измерения вращательной диффузии молекул давно используется метод деполяризации флуоресценции. Вращение некоторых зондов в мембранах часто сравнивают с их вращением в маслах с известной вязкостью, при этом пользуются понятием «микровязкость», которую измеряют в пуазах. Термин «микровязкость» означает, что зонд «чувствует» величину вязкости только в своем непосредственном окружении, т. е. этот параметр не является макроскопическим. С точки зрения молекулярного движения подобные измерения не имеют строгого количественного смысла, но, как и в случае со спиновыми метками, полученные результаты позволяют создать ценную качественную картину.
В табл. 1 приведены структурные формулы некоторых зондов, используемых при измерении текучести мембран. Чаще всего применяют ДФГ, поскольку он с легкостью накапливается в мембранах, обладает интенсивной флуоресценцией, по-видимому, не связывается с белками и чувствителен к физическому состоянию мембран. Однако временные измерения свидетельствуют о некой гетерогенности неизвестной природы; кроме того, имеются разногласия по поводу ориентации зонда в бислое и характера его подвижности.
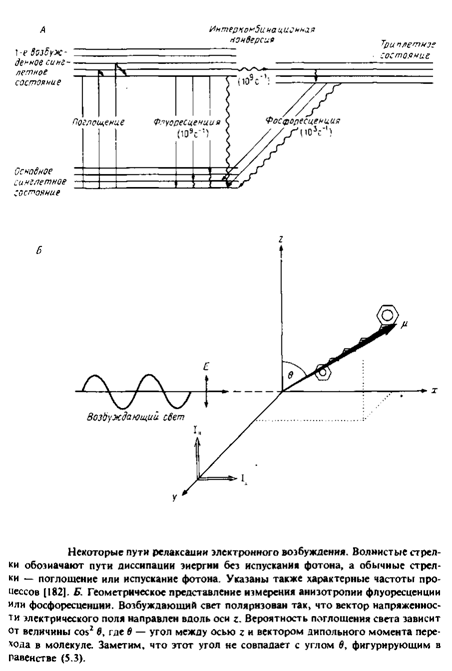
Рис. 4 иллюстрирует принципы, лежащие в основе применения флуоресценции и фосфоресценции для анализа вращения молекул. Молекула, поглотившая фотон, за время 10" 11 с переходит в возбужденное синглетное состояние Si. Термин «синглетное» указывает на то, что спин электрона при поглощении фотона не изменяется. Обычно возвращение в основное, невозбужденное состояние происходит за время - 10"8 с. Причинами такого перехода являются столкновение с соседними молекулами, диссипация энергии в виде тепла или испускание фотона. Характерное время перехода популяции молекул из синглетного возбужденного состояния в основное равно тт. Некоторые молекулы с большой вероятностью переходят из синглетного возбужденного состояния в триплетное, и этот переход сопровождается изменением спина электрона. Испускание фотона при переходе молекулы из этого состояния в основное называется фосфоресценцией. Время жизни молекулы в триплетном состоянии относительно велико, что связано с изменением спинового состояния электрона при испускании фотона.
Рассмотрим принцип метода деполяризации. Свет, поляризованный вдоль оси z, будет предпочтительно возбуждать молекулы, у которых дипольный момент перехода имеет такую же ориентацию. Вероятность возбуждения пропорциональна cos2 в, где в — угол между дипольиым моментом перехода и вектором напряженности электрического поля. Такая же зависимость от cos2 в имеет место и для процесса испускания фотона, поэтому молекула, у которой дипольный момент перехода ориентирован вдоль оси z, будет испускать свет с такой же поляризацией.
Испускаемый свет анализируют с помощью поляризаторов, определяя интенсивность компоненты, параллельной и перпендикулярной направлению поляризации возбуждающего света.
ИНТЕРЕСНОЕ
© 2009 Все права защищены. |