 | |
МЕНЮ
- Главная
- Языкознание филология
- Финансовые науки
- Управленческие науки
- Товароведение
- Технология
- Теплотехника
- Теория организации
- Теория государства и права
- Таможенная система
- Схемотехника
- Строительство
- Страхование
- Статистика
- Религия и мифология
- Психология и педагогика
- Промышленность производство
- Медицинские науки
- Медицина
- Краеведение и этнография
- Компьютерные науки
- История
- Искусство и культура
- Информатика
- Инвестиции
- Издательское дело и полиграфия
- Зоология
- Журналистика
- Естествознание
- Деньги и кредит
- Делопроизводство
- Гражданское право и процесс
- Государство и право
- Геополитика
- Геология
- Геодезия
- География
- Военная кафедра
- Ветеринария
- Валютные отношения
- Бухгалтерский учет и аудит
- Ботаника и сельское хоз-во
- Биржевое дело
- Биология и химия
- Биология
- Безопасность жизнедеятельности
- Банковское дело
- Астрономия
- Астрология
- Архитектура
- Арбитражный процесс
- Административное право
- Авиация и космонавтика
- Карта сайта
Реферат: Действие низкой температуры на растения
Реферат: Действие низкой температуры на растения
Действие низкой температуры на растения
За более чем столетний период изучения действия на клетки растений низких положительных и отрицательных температур знания о вызываемых ими изменениях в метаболизме растительных клеток, а также о свойствах клеток растений, определяющих их морозостойкость, значительно углубились и расширились.
В настоящее время считается, что основными причинами, вызывающими гибель растений от холода, являются или непосредственное действие низких температур на клетки, не связанное с образованием льда в тканях, или же образование льда в тканях снаружи клеток либо внутри них. С одной стороны, было установлено, что внутриклеточное замерзание воды всегда приводит к их немедленно гибели, во всяком случае, в лабораторных условиях, имитирующих природные. С другой стороны, все растения, зимующие в условиях умеренного климата, переносят внеклеточное замерзание значительных количеств воды.
Мгновенное и необратимое повреждение клеток при образовании внутриклеточного льда указывает на физическую природу процесса: вероятнее всего происходит разрушение мембран клетки растущими в протоплазме кристаллами льда. В тоже время, рассматривая причины вымерзания растений, Г.А. Самыгин отмечает, что в природных условиях очень редко имеет место внутриклеточное образование льда, поскольку оно возможно только при очень быстром снижении температуры, достигающим 10–120С в час. В естественных же условиях температура воздуха снижается со скоростью 1–20С в час или даже еще медленнее. В этом случае происходит внеклеточное образование льда.
Повреждения при образовании льда вне клеток вызываются двумя основными причинами: обезвоживанием протопластов и механическими повреждениями обезвоженной протоплазмы. У закаленных растений преимущественно причиной гибели являются механические повреждения, у незакаленных – обезвоживание протоплазмы.
Обезвоживание растительной клетки имеет ряд опасных для нее последствий, которые могут привести ее к гибели. Это, во-первых, повышение концентрации растворенных веществ, и, прежде всего, солей; во-вторых, изменение рН внутриклеточных растворов; в третьих, образование ковалентных связей между макромолекулами; в четвертых, конформационные изменения структуры макромолекул из-за снижения содержания стабилизирующих их молекул воды; в-пятых, нарушение структуры мембран, и наконец, в шестых, это повреждение структуры протоплазмы при обратном поглощении воды.
Особенно чувствительными структурами клетки, легко повреждающимися под действием гипотермии, являются клеточные мембраны. Функциональная устойчивость липидосодержащих протоплазматических структур, таких как плазмалемма и мембраны хлоропластов и митохондрий, легко нарушается под действием экстремальных температур. В частности, рядом авторов было показано, что замораживание вызывает необратимое подавление окислительного фосфорилирования в митохондриях. По их мнению, основной причиной повреждения клетки морозом является нарушение структуры мембран и потеря ими осмотических свойств. По мнению других авторов, важнейший фактор повреждения мембран при замораживании – их обезвоживание. Удаление воды из мембран при замораживании нарушает равновесие между системами белок – липид и белок – вода, в результате чего возникают структурные перестройки молекулярных слоев и изменяются свойства мембран. Обособление белков от липидов в мембране приводит к полному ее разрушению и в дальнейшем мембранные липиды могут стать субстратом для окисления. Наличие лишь незначительного числа нарушений мембранной структуры вызывает утечку протонов и разобщение окисления и фосфорилирования. Возможно повреждение мембран и без образования льда, когда низкая температура вызывает затвердение липидной части мембраны и нарушение ее структуры и функций.
Впоследствии многочисленные исследования показали сильную зависимость состава мембран растительной клетки от температуры роста растения. В частности, показано, что низкие температуры индуцируют в растениях разных видов накопление фосфолипидов и повышение ненасыщенности липидов. При этом были отмечены различия в содержании фосфолипидов у высоко- и низкохолодостойких сортов растений. В частности, такие различия были отмечены у высоко- и низкохолодостойких сортов люцерны, а также различающихся по холодоустойчивости сортов озимой пшеницы, при этом наибольшие отличия в фосфолипидном составе листьев высоко- и низкохолодостойких сортов озимой пшеницы были отмечены после прохождения растением процесса низкотемпературно адаптации. Наиболее важным фактором в этом случае было возрастание доли фосфатидилхолина в составе мембран. Сходные результаты были получены и при изучении проростков озимой пшеницы. У проростков, выращенных при 20С, было обнаружено значительно более высокое содержание фосфолипидов, чем у проростков, выращенных при оптимальных температурах. У растений озимой пшеницы наблюдались также генотипические особенности содержания фосфолипидов в узлах кущения. Содержание как суммарных фосфолипидов, так и их отдельных фракций, таких как фосфатидилхолина и фосфатидилэтаноламина, было выше у морозостойкого сорта. Аналогичные результаты были получены и при изучении липидного состава корне озимой ржи во время роста при пониженных температурах.
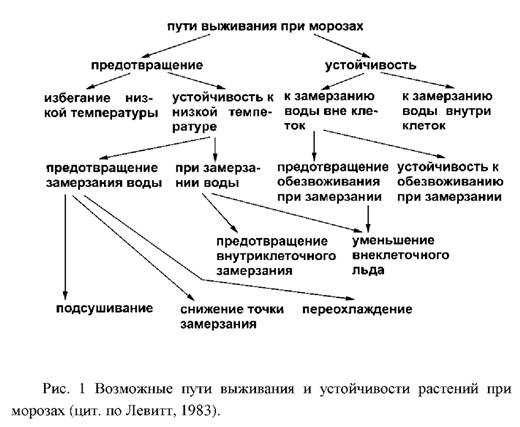
Для того, чтобы растения выживали после действия гипотермии, необходимо или предотвращение повреждений от действия внеклеточного или внутриклеточного замерзания, или ликвидация таких повреждений. Это свойство растений, определяемое как морозоустойчивость, должно включать в себя как предотвращение воздействия повреждающих факторов, так и устойчивость к их действию. Предотвращение воздействия повреждающих факторов возможно следующими путями:
– предотвращение воздействия низко температуры;
– предотвращение замерзания путем высушивания;
– предотвращение замерзания нормально гидратированных клеток может осуществляться путем накопления растворимых веществ;
– предотвращение замерзания вследствие переохлаждения обеспечивает выживание определенных тканей у многих древесных культур;
– предотвращение образования льда внутри клеток у растений обеспечивается различными способами и сохраняет жизнеспособность растений лишь в том случае, если они устойчивы к внеклеточному замерзанию;
– уменьшение количества льда при замерзании воды вне клеток идентично предотвращению обезвоживания при замерзании.
В большинстве случаев для выживания растений недостаточно предотвращения повреждений, а необходима еще и устойчивость к замораживанию некоторых тканей и, вследствие этого, их обезвоживанию. Растение может переносить обезвоживание тканей морозом двумя путями: во-первых, путем предотвращения летального обезвоживания при замерзании посредством действия накопленных растворимых веществ и, во-вторых, повышением устойчивости растений к обезвоживанию при замерзании, обусловливающей высокую морозостойкость тех видов, у которых не наблюдается корреляция между степенью морозоустойчивости и накоплением растворимых соединений.
В отличие от древесных растений, травянистые не входят в конце лета в состояние глубокого покоя. Их ткани, несмотря на вынужденную приостановку роста, сохраняют способность к нему на протяжении все зимы. В то же время, несмотря на то, что неблагоприятные температурные условия вызывают в их клетках ряд последовательных изменений: снижение водного потенциала, нарушение метаболизма и др. У видов, способных к закаливанию, надземные ткани переносят температуру до -20…– 25°С после предварительного воздействия закаливающих условий.
Закаливание озимых растений происходит в три фазы, которые связаны с различными температурными условиями:
– первая фаза закаливания индуцируется снижением температуры до 2 – 5°С, в результате чего исходная морозостойкость повышается на 4–5°С;
– вторая фаза связана с небольшими морозами и в естественных условиях проходит только в том случае, если температура воздуха снижается ниже 0°С;
– третья фаза закаливания может совпадать со второй и зависит от продолжительных морозов, вызывающих обезвоживание клеток.
При изучении процессов закаливания различными исследователями было установлено, что во время первой фазы закаливания в растениях происходят изменения метаболических процессов, таких как гидролиз крахмала и накопление редуцирующих сахаров и водорастворимых белков. Другими метаболическими изменениями, которые тоже являются специфическим ответом растительных тканей на понижение температуры среды, являются превращения липидов и фосфолипидов. Установлено, что низкие положительные температуры повышают ненасыщенность жирных кислот. Было также показано, что содержание фосфолипидов в тканях растений пшеницы возрастало при низкотемпературном закаливании.
Существует ряд доказательств того, что накопление в тканях растений в результате воздействия низких положительных температур некоторых веществ происходит не только вследствие уменьшения их утилизации в ходе ростовых процессов ввиду их приостановки, но и в результате усиления их новообразования. Увеличение ненасыщенности жирных кислот фосфолипидов в условиях пониженных температур, как было показано, тоже в значительно степени связано с их синтезом. Все эти данные свидетельствуют о том, что на первой фазе закаливания у травянистых растений происходит предпочтительны синтез некоторых метаболитов.
Что касается отрицательных температур, которые необходимы для прохождения второй фазы закаливания, то предполагается, что их роль в процессе закаливания не сводится только к физическому действию. По-видимому, она также связана с некоторыми метаболическими процессами, протекающими при этих температурах.
Поскольку прекращение роста растений является необходимым условием прохождения первой фазы закаливания, то метаболические изменения, происходящие в это время, могут быть вызваны изменением гормонального и энергетического баланса.
Принимая во внимание тот факт, что внеклеточное замерзание предотвращает образование внутриклеточного льда, но вызывает при этом обезвоживание макромолекулярных структур клетки, предполагается, что процесс закаливания включает в себя следующие механизмы:
– усиление оттока воды из клетки через мембраны;
– защита клеточных компонентов от действия обезвоживания. Обеспечение оттока воды через мембраны может обеспечиваться путем повышения ненасыщенности липидов. Изменения содержания фосфолипидов также влияют на свойства мембран и повышают их проницаемость для воды, а быстрое снижение содержания фосфолипидов при замораживании вызывает усиленны отток воды в межклетники и защищает клетку от внутриклеточного льдообразования.
При действии мороза повреждения, вызываемые обезвоживанием у закаленных растений, могут быть предотвращены следующими путями:
– структурными и конформационными изменениями компонентов клетки, которые они претерпевают в процессе закаливания;
– защитой компонентов клетки от обезвоживания взаимодействием с низкомолекулярными веществами.
Хотя и установлено, что под действием гипотермии у травянистых растений происходят изменения активности некоторых ферментов, имеется относительно немного экспериментальных доказательств трансформации белковых макромолекул, которые бы вели к повышению их устойчивости к низко температуре. В то же время показано большое значение происходящего во время закаливания растений торможения роста для использования белков, синтезированных в процессе закаливания, на структурную и функциональную реорганизацию клеток.
Исследование процессов холодовой адаптации древесных растений на молекулярном уровне ограничивается наложением на процессы развития холодоустойчивости событий, связанных со входом растения в состояние покоя. R.A. Salzman с соавторами, используя в качестве объекта исследования виноград, создали систему, в которой развитие состояния покоя могло быть индуцировано отдельно от холодовой акклиматизации. С использованием это системы было охарактеризовано дифференциальное накопление ряда белков в почках винограда во время реализации программы нормального входа в состояние покоя совместно с холодовой акклиматизацией и в почках, которые входили только в состояние покоя. Было установлено, что белок с молекулярно массой 47 кДа накапливался в почках винограда во время входа в состояние покоя без холодовой акклиматизации до уровня содержания белка, обнаруженного в находящихся в состоянии покоя и закаленных почках, но не накапливался в закаленных почках, не вошедших в состояние покоя. В то же время 27 кДа LEA-подобный белок накапливался только в закаленных почках. Следовательно, 47 кДа гликопротеин является связанным с состоянием покоя, но не связанным с развитием холодовой акклиматизации, в то время как 27 кДа LEA-подобны белок, по-видимому, более специфичен для холодового закаливания.
Большое значение в регуляции холодо- и морозоустойчивости растений играет абсцизовая кислота. Установлено, что при закаливании растений содержание эндогенно абсцизовой кислоты значительно возрастает, в частности, при закаливании способного к холодовой адаптации вида картофеля Solanum commersonii содержание эндогенной абсцизовой кислоты возрастало в 2,5 раза. У мутанта Arabidopsis thaliana, имеющего низкий уровень содержания эндогенно абсцизовой кислоты, по сравнению с диким типом отсутствовала или была резко снижена способность к холодовой адаптации. В то же время обработка этого мутанта экзогенно абсцизовой кислотой приводила к появлению эффекта адаптации растений к холоду. При этом была показана взаимосвязь между экспрессией регулируемых холодом и регулируемых абсцизовой кислотой генов.
Аналогичные результаты были получены и при изучении других видов растений. Было проведено определение морозоустойчивости побегов, корней и тканей эпикотиля гороха сорта Alaska двух генотипов: дефицитного по содержанию абсцизовой кислоты мутанта «wil» и его дикого типа при различных типах стресса. В ходе исследований спектры белков изучались при помощи двумерного SDS-PAGE электрофореза. При этом было установлено, что холодовая обработка индуцировала образование семи белков в побегах, трех – в эпикотиле и двух – в корнях гороха. В тканях побегов пять из семи новых белков накапливались также в ответ на обработку абсцизовой кислотой. Полипептид с молекулярной массой 24 кДа продуцировался и в мутантных, и в «диких» проростках и тканях эпикотиля только после холодовой обработки.
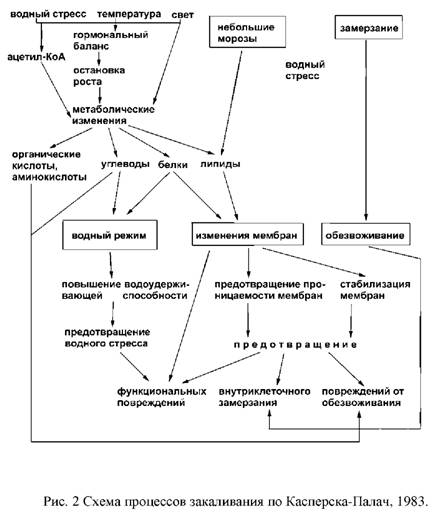
Таким образом, существенным этапом перехода от стрессовых к адаптационным реакциям является изменение экспрессии генов, выражающееся в ингибировании активных генов, в норме контролирующих рост, развитие и фотосинтез. При этом активируется система генов контроля за устойчивостью: происходит синтез новых белков, специфических адаптогенов и стресс-протекторов. Завершается эта перестройка структурными изменениями в организме растения.
Успешное зимнее выживание вечнозеленых травянистых растений, подобных белому клеверу, зависит от соответствующе синхронизации процессов как закаливания, так и раззакаливания. Изучение регулирования этих процессов было проведено у двух сортов белого клевера «AberCrest» и «AberHerald» и двух его норвежских экотипах. Для проведения закаливания и раззакаливания растения экспонировались при контролируемых температурных условиях. Низкотемпературное закаливание столонов проводилось путем программируемого снижения температуры со скоростью 3°C в час. Во время эксперимента анализировались содержание крахмала, растворимых сахаров и растворимых аминокислот в столонах. Сорта AberCrest и AberHerald, происходящие из Великобритании и выбранные для контроля скорости роста при низко температуре и степени зимнего закаливания, были значительно менее устойчивы, чем норвежские популяции. Степень раззакаливания растени увеличивалась с повышением температуры. В условиях действия низко температуры северный экотип из Bodo был более устойчив к раззакаливанию, чем AberHerald. Тем не менее, при 18°C абсолютны уровень раззакаливания у растений экотипа Bodo был в два раза выше, чем у растений AberHerald. Удлинение столонов в растениях AberHerald начиналось во время раззакаливания при более низких температурах, чем в растениях экотипа Bodo. Содержание общих растворимых сахаров, сахарозы и аминокислот пролина и аргинина было значительно выше в закаленных растениях экотипа Bodo, чем в растениях сорта AberHerald. Уровень сахарозы уменьшался в течение раззакаливания. Корреляция между содержанием сахарозы и LT50 в течение этого процесса была статистически достоверно.
Установлено, что одним из криопротекторов в растениях является глицинбетаин. Это вещество накапливается в хлоропластах определенных солеустойчивых растений при солевом или холодовом стрессах. Ген codA для холиноксидазы, преобразовывающей холин в глицинбетаин, был клонирован в почвенной бактерии Arthrobacter globiformis. Трансформация Arabidopsis thaliana с клонированным геном codA под управлением 35S промотора мозаичного вируса цветной капусты позволила растению накапливать глицинбетаин и увеличить устойчивость к солевому и холодовому стрессам. Значительная часть семян трансформированных растений хорошо прорастала в 300 мл NaCl, в то время как семена растений дикого генотипа в данных условиях не прорастали. В растворе NaCl трансформированные растения хорошо росли, в то время как растения дикого типа не были способны расти в данных условиях. Трансформированные растения были способны переносить концентрацию 200 мл NaCl, которая была летально для растений дикого типа. После того, как растения были инкубированы в течение двух дне в растворе с повышенно концентрацией NaCl, активность фотосистемы II растений дикого типа была почти полностью подавлена, в то время как в трансформированных растениях она составляла более 50% от исходного уровня. После обработки растений низко температурой на свету в листьях дикого типа наблюдались симптомы хлороза, в то время как у трансформированных растений они отсутствовали. Эти наблюдения показывают, что генетическая трансформация, позволяющая накапливать глицинбетаин Arabidopsis thaliana, увеличивает способность растения переносить солевой и низкотемпературный стрессы.
Было изучено влияние низко температуры на биосинтез полиаминов в листьях, стеблях и корнях разновидностей озимой пшеницы с различно морозоустойчивостью. Оказалось, что в этих условиях происходит заметное накопление полиаминов. Кроме того, обнаружен эффект 5A и 7A хромосом пшеницы, содержащих основные гены, ответственные за морозоустойчивость, на синтез полиаминов, происходящий в различных частях проростков в течение длительных периодов холодовой обработки.
Для того, чтобы определить, будет ли система in vitro подходить для изучения зимнего покоя и закаливания у древесных растений, культивируемые in vitro растения ирги ольхолистно были подвергнуты различной гормональной обработке, индукции зимнего покоя и акклиматизации к низким температурам. Низкие температуры вызвали значительное повышение уровня холодоустойчивости растений, но она не приблизилась к уровню холодоустойчивости полностью закаленных почек, которые способны переносить температуру жидкого азота. Контрольные растения в данном эксперименте полностью гибли при -50C. Значительны уровень закаленности был достигнут при действии низких температур и при коротком, и при длинном дне. Предварительная обработка низко температурой при типичном для древесных растений режиме короткого фотопериода незначительно увеличила степень акклиматизации в этих растениях. Присутствие или отсутствие фитогормонов в среде имеет выраженное влияние на холодовую акклиматизацию растений. Безгормональная среда после 2 недель обработки увеличила холодоустойчивость до -10.50C. Добавление в среду абсцизовой кислоты увеличило уровень холодовой закаленности, в то время как добавление бензиламинопурина к безгормональной среде уменьшило закаленность до -5.30C. Сочетание обработок бензиламинопурином и абсцизовой кислотой изменяло значения LT50 до промежуточных между индивидуальными обработками любым гормоном. Напротив, x-нафталенацетиловая кислота не снижала индуцированную абсцизовой кислотой закаленность. Обработка абсцизовой кислотой, как таковая, не была способна закалить растения до уровня, достигаемого при акклиматизирующем действии низко температуры. Далее, абсцизовая кислота не могла поддерживать уровень закаленности после холодово акклиматизации и растения деакклиматизировались до 90C на среде бензиламинопурин + абсцизовая кислота. Культивирование в не значительно увеличивало закаленность к холоду в растениях, но впоследствии растения раззакаливались до –50C.