 | |
МЕНЮ
- Главная
- Языкознание филология
- Финансовые науки
- Управленческие науки
- Товароведение
- Технология
- Теплотехника
- Теория организации
- Теория государства и права
- Таможенная система
- Схемотехника
- Строительство
- Страхование
- Статистика
- Религия и мифология
- Психология и педагогика
- Промышленность производство
- Медицинские науки
- Медицина
- Краеведение и этнография
- Компьютерные науки
- История
- Искусство и культура
- Информатика
- Инвестиции
- Издательское дело и полиграфия
- Зоология
- Журналистика
- Естествознание
- Деньги и кредит
- Делопроизводство
- Гражданское право и процесс
- Государство и право
- Геополитика
- Геология
- Геодезия
- География
- Военная кафедра
- Ветеринария
- Валютные отношения
- Бухгалтерский учет и аудит
- Ботаника и сельское хоз-во
- Биржевое дело
- Биология и химия
- Биология
- Безопасность жизнедеятельности
- Банковское дело
- Астрономия
- Астрология
- Архитектура
- Арбитражный процесс
- Административное право
- Авиация и космонавтика
- Карта сайта
Реферат: Функциональная организация мозга
Реферат: Функциональная организация мозга
Московский гуманитарно-экономический институт
Тверской филиал
Кафедра прикладной психологии
Реферат по дисциплине
«Физиология высшей нервной деятельности и сенсорных систем»
Тема: «Функциональная организация мозга».
Тверь 2009
Содержание
Введение
1. Три основных функциональных блока мозга
1.1 Блок приема и переработки сенсорной информации
1.2 Блок модуляции, активации нервной системы
1.3 Блок программирования, запуска и контроля поведенческих актов
2. Взаимодействие трех основных функциональных блоков мозга
Заключение
Список литературы
Введение
Открытие И.П. Павловым анализаторов и создание учения об условных рефлексах, в основе которого лежал объективный анализ динамики нервных процессов, послужило основой для развития современных материалистических представлений о динамической локализации мозговых функций - целостном и одновременно дифференцированном вовлечении мозга в любую из форм его активности.
Предложенный И.П. Павловым объективный условно рефлекторный метод исследования позволил наиболее адекватно подойти к экспериментальному решению проблемы функциональной организации мозга. И.П. Павлов развил и экспериментально обосновал представления об анализаторных системах, где каждый анализатор есть определенная анатомически локализованная структура от периферических рецепторных образований до проекционных зон коры головного мозга. Он предположил, что кроме локальных проекционных зон коры, выступающих в качестве «ядра коркового конца анализатора» (или проекционных зон коры), существуют периферические зоны представительства каждого анализатора, так называемые «зоны рассеянных элементов». В силу такой структурной организации все анализаторы, включая и двигательный анализатор, своими периферическими (корковыми) зонами перекрываются и образуют вторичные проекционные зоны коры, которые И.П. Павлов уже тогда рассматривал как «ассоциативный центры» мозга, составляющие основу для динамического взаимодействия всех анализаторных систем.
С позиций системной организации функций в деятельности мозга выделяют различные функциональные системы и подсистемы. Классический вариант интегративной деятельности мозга может быть представлен в виде взаимодействия трех основных функциональных блоков:
1) блок приема и переработки сенсорной информации - сенсорные системы (анализаторы);
2) блок модуляции, активации нервной системы - модулирующие системы (лимбико-ретикулярные системы) мозга;
3) блок программирования, запуска и контроля поведенческих актов - моторные системы (двигательный анализатор).
1. Три основных функциональных блока мозга
1.1 Блок приема и переработки сенсорной информации
Первый функциональный блок составляют анализаторы, или сенсорные системы. Анализаторы выполняют функцию приема и переработки сигналов внешней и внутренней среды организма. Каждый анализатор настроен на определенную модальность сигнала и обеспечивает описание всей совокупности признаков воспринимаемых раздражителей.
Анализатор - это многоуровневая система с иерархическим принципом ее конструкции. Основанием анализатора служит рецепторная поверхность, а вершиной - проекционные зоны коры. Каждый уровень этой морфологически упорядоченно организованной конструкции представляет собой совокупность клеток, аксоны которых идут на следующий уровень (исключение составляет верхний уровень, аксоны которого выходят за пределы данного анализатора). Взаимоотношения между последовательными уровнями анализаторов построены по принципу «дивергенции - конвергенции». Чем выше нейронный уровень анализаторной системы, тем большее число нейронов он включает. На всех уровнях анализатора сохраняется принцип топической проекции рецепторов. Принцип многократной рецептотопической проекции способствует осуществлению множественной и параллельной переработки (анализу и синтезу) рецепторных потенциалов («узоров возбуждений»), возникающих под действием раздражителей.
Нейрон, стоящий на выходе рецептивного поля, может выделять один признак раздражителя (простые детекторы) или комплекс его свойств (сложные детекторы). Детекторные свойства нейрона обусловливаются структурной организацией его рецептивного поля. Нейроны-детекторы более высокого порядка образуются в результате конвергенции нейронов-детекторов низшего (более элементарного) уровня. Нейроны-детекторы сложных свойств формируют детекторы «сверхсложных» комплексов. Высший уровень иерархической организации детекторов достигается в проекционных зонах и ассоциативных областях коры мозга.
Проекционные зоны анализаторных систем занимают наружную (конвекситальную) поверхность новой коры задних отделов мозга. Сюда входят зрительная (затылочная), слуховая (височная) и общечувствительная (теменная) области коры. В корковый отдел этого функционального блока включается также представительство вкусовой, обонятельной, висцеральной чувствительности.
Первичные проекционные зоны коры состоят главным образом из нейронов 4-го афферентного слоя, для которых характерна четкая топическая организация. Значительная часть этих нейронов обладает высочайшей специфичностью. Нейроны зрительных аппаратов коры реагируют только на узкоспециальные свойства зрительных раздражителей (оттенки цвета, характер линий, направление движения). Однако следует отметить, что в первичные зоны отдельных областей коры включены также нейроны мультимодального типа, реагирующие на несколько видов раздражителей.
Вторичные проекционные зоны коры располагаются вокруг первичных зон, как бы надстраиваясь над ними. В этих зонах 4-й афферентный слой уступает ведущее место 2-му и 3-му слоям клеток. Для этих нейронов характерно детектирование сложных признаков раздражителей, однако при этом сохраняется модальная специфичность, соответствующая нейронам первичных зон. Поэтому предполагается, что усложнение детекторных селективных свойств нейронов вторичных зон может происходить путем конвергенции на них нейронов первичных зон. В первичной зрительной коре (17-е поле Бродмана) содержатся в основном нейроны-детекторы простых признаков предметного зрения (детекторы ориентации линий, полосы, контраста и т. п.), а во вторичных зонах (18-е и 19-е поля Бродмана) появляются детекторы более сложных элементов контура: края, ограниченной длины линий, углов с различной ориентацией и др. Первичные (проекционные) зоны слуховой (височной) коры представлены 41-м полем Бродмана (рис. 1), нейроны которого модально специфичны и реагируют на различные свойства звуковых раздражителей. Как и первичное зрительное поле, эти первичные отделы слуховой коры имеют четкую рецептотопию. Над аппаратами первичной слуховой коры надстроены вторичные зоны слуховой коры, расположенные во внешних отделах височной области (22-е и частично 21-е поля Бродмана). Они также состоят преимущественно из мощно развитого 2-го и 3-го слоя клеток, реагирующих избирательно одновременно на несколько частот и интенсивностей: звукового раздражителя.
Рис. 1. Карта цитоархитектонических полей коры головного мозга. Конвекситальная поверхность коры больших полушарий: а — первичные поля; б — вторичные поля; в — третичные поля
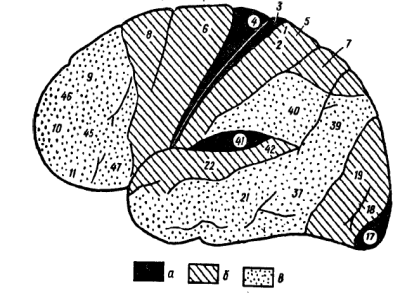
Наконец, тот же принцип функциональной организации сохраняется и в общечувствительной (теменной) коре. Основой и здесь являются первичные или проекционные зоны (3-, 1- и 2-е поля Бродмана), толща которых также преимущественно состоит из обладающих модальной специфичностью нейронов 4-го слоя, а топография отличается четкой соматотопической проекцией отдельных сегментов тела. Вследствие чего раздражение верхних участков этой зоны вызывает появление кожных ощущений в нижних конечностях, средних участков - в верхних конечностях контрлатеральной стороны, а раздражение пунктов нижнего пояса этой зоны - соответствующие ощущения в контрлатеральных отделах лица, губ и языка. Над первичными зонами располагаются вторичные зоны общечувствительной (теменной) коры (5-е и частично 40-е поле Бродмана), состоящие преимущественно тоже из нейронов 2-го и 3-го слоев, и их раздражение приводит к возникновению более комплексных форм кожной и кинестетической чувствительности (см. рис. 1).
Таким образом, основные, модально-специфические зоны анализаторов мозга построены по единому принципу иерархической структурной и функциональной организации. Первичные и вторичные зоны, согласно И.П. Павлову, составляют центральную часть, или ядро, анализатора в коре, нейроны которого характеризуются избирательной настройкой на определенный набор параметров раздражителя и обеспечивают механизмы тонкого анализа и дифференцировки раздражителей. Взаимодействие первичных и вторичных зон носит сложный, неоднозначный характер и в условиях нормальной деятельности обусловливает согласованное содружество процессов возбуждения и торможения, которое закрепляет макро- и микроструктуру нервной сети, занятой анализом афферентного потока в первичных проекционных сенсорных полях. Это создает основу для динамического межанализаторного взаимодействия, осуществляемого в ассоциативных зонах коры.
Ассоциативные области (третичные зоны) коры являются новым уровнем интеграции: они занимают 2-й и 3-й клеточные (ассоциативные) слои коры, на которых протекает встреча мощных афферентных потоков, как одномодальных, разномодальных, так и неспецифических. Подавляющее большинство ассоциативных нейронов отвечает на обобщенные признаки стимулов - на количество элементов, пространственное положение, отношения между элементами и пр.
Конвергенция разномодальной информации необходима для целостного восприятия, для формирования «сенсорной модели мира», которая возникает в результате сенсорного обучения.
Ассоциативные зоны расположены на границе затылочной, височной и заднетеменной коры. Работа этих отделов коры мозга необходима не только для успешного синтеза и дифференцировки избирательного различения воспринимаемых человеком раздражителей, но и для перехода к уровню их символизации - для оперирования значениями слов и использования их для отвлеченного мышления.
Клинические наблюдения различных очаговых поражений третичных зон мозга человека накопили большой материал о взаимосвязи ассоциативных областей с различными функциональными расстройствами. Известно, что поражения лобно-височно-теменной области, так называемых речевых зон (имеется в виду левое полушарие), связаны с возникновением афазии (расстройства речи). При поражении нижневисочной области наблюдают предметную агнозию (нарушение процесса узнавания), теменных областей или угловой извилины теменной доли — развитие оптико-пространетвенной агнозии, при поражении левой височной доли обнаруживается цветовая агнозия и т. д. Следует отметить, что локальные поражения ассоциативных зон коры могут быть связаны как с относительно элементарными сенсорными расстройствами, так и с нарушениями сложных форм восприятия.
1.2 Блок модуляции, активации нервной системы
Блок модулирующих систем мозга регулирует тонус коры и подкорковых образований, оптимизирует уровень бодрствования в отношении выполняемой деятельности и обусловливает адекватный выбор поведения в соответствии с актуализированной потребностью. Только в условиях оптимального бодрствования человек может наилучшим образом принимать и перерабатывать информацию, вызывать в памяти нужные избирательные системы связей, программировать деятельность, осуществлять контроль над ней.
В условиях оптимальной возбудимости коры нервные процессы характеризуются известной концентрированностью, уравновешенностью возбуждения и торможения, способностью к дифференцировке и, наконец, высокой подвижностью нервных процессов, которые обусловливают протекание каждой организованной целенаправленности деятельности.
Аппаратом, выполняющим роль регулятора уровня бодрствования, а также осуществляющим избирательную модуляцию и актуализацию приоритета той или иной функции, является модулирующая система мозга. Ее часто называют лимбико-ретикулярный комплекс или восходящая активирующая система. К нервным образованиям этого аппарата относятся лимбическая и неспецифическая система мозга с ее активирующими и инактивирующими структурами. Среди активирующих образований прежде всего выделяют ретикулярную формацию среднего мозга, задний гипоталамус, синее пятно в нижних отделах ствола мозга. К инактивирующим структурам относят преоптическую область гипоталамуса, ядра шва в стволе мозга, фронтальную кору.
Важнейшей частью модулирующего блока мозга является активирующая ретикулярная формация. Филогенетически ретикулярная формация мозга представляет наиболее древнее морфологическое образование. В ретикулярной формации выделяют более или менее компактные и ограниченные клеточные скопления - ядра, отличающиеся различными морфологическими особенностями. В связи с этим одни авторы рассматривают ретикулярную формацию как диффузное вытянутое в длину единое образование, другие считают ее комплексом, состоящим из многих дифференцированных ядер с различной структурой и функциями. Латерально (с боков) ретикулярная формация окружена сенсорными путями. Таким образом, волокна ретикулярной формации окружены слоем сенсорных путей, которые к ней образуют множество коллатералей.
Функциональное назначение ретикулярной формации долго оставалось неизвестным. Электрофизиологические исследования выявили исключительную роль ретикулярной формации в интегративной деятельности мозга. Это открытие было сделано в 1949 г. Г. Мэгуном и Г. Моруцци. Путем стимуляции через электроды, вживленные в стволовой отдел мозга (на уровне среднего мозга), им удалось получить реакцию пробуждения спящего животного. Эту стволовую систему мозга Г. Мэгун назвал восходящей активирующей системой мозга.
Волокна ретикулярной формации, направляясь вверх, образуют модулирующие «входы» (как правило, аксодендритные синапсы) в выше расположенных мозговых образованиях, включая старую и новую кору. От старой и новой коры берут начало нисходящие волокна, которые идут в обратном направлении к структурам гипоталамуса, среднего мозга и к более низким уровням мозгового ствола. Через нисходящие системы связей все нижележащие образования оказываются под управлением и контролем тех программ, которые возникают в коре головного мозга и для выполнения которых требуется модуляция активности и модификация состояний бодрствования. Таким образом, блок активации с его восходящими и нисходящими влияниями работает (по принципу обратной связи) как единый саморегулирующий аппарат, который обеспечивает изменение тонуса коры, и вместе с тем сам находится под его контролем. Этот аппарат используется для пластичного приспособления организма к условиям среды. Он содержит в своей основе по крайней мере два источника активации: внутренний и внешний. Первый связан с обменными процессами, обеспечивающими внутреннее равновесие организма, второй — с воздействием внешней среды. Первым источником активации является внутренняя активность самого организма, или потребности. Любые отклонения от жизненно важных «констант» в результате изменения нервных или гуморальных влияний или вследствие избирательного возбуждения различных отделов мозга приводят к выборочному «включению» определенных органов и процессов, совокупная работа которых обеспечивает достижение оптимального состояния для данного вида деятельности организма.
Наиболее простые формы внутренней активности связаны с дыхательными и пищеварительными процессами, процессами внутренней секреции и другими, включенными в гомеостатический механизм саморегуляции, который устраняет нарушение во внутренней среде организма за счет своих резервов. Более сложные формы этого вида активации организованы в структуру врожденного поведения, направленного на удовлетворение определенной потребности. Естественно, для того чтобы обеспечить механизм инстинктивной регуляции поведения, необходима весьма избирательная и специфическая активация. Такая специфическая активация может быть функцией лимбической системы мозга, в которой важная роль принадлежит гипоталамусу.
Гипоталамус - часть промежуточного мозга, содержит десятки высоко дифференцированных ядер, обладающих обширной и разносторонней системой связей. Гипоталамус координирует внутренние потребности организма с его внешним поведением, направленным на достижение приспособительного эффекта. Гипоталамус входит в состав потребностно-мотивационной системы, являясь ее главной исполнительной структурой. При этом он не просто участвует в регуляции отдельных жизненно важных функций (голода, жажды, полового влечения, активной и пассивной обороны), а осуществляет их объединение в сложные комплексы или системы. В зависимости от характера нервной и гуморальной сигнализации, собирающейся в гипоталамусе, в нем или накапливается, или тормозится мотивационное возбуждение, определяющее внешнее поведение.
Второй источник активации связан с воздействием раздражителей внешней среды. Ограничение контакта с внешней средой приводит к значительному снижению тонуса (возбудимости) коры мозга. В условиях резкого ограничения сенсорной информации у человека могут возникать галлюцинации, которые в какой-то мере компенсируют дефицит сенсорного возбуждения.
Часть непрерывного потока сенсорных сигналов, поставляемых в кору специфическими (анализаторными) системами, по коллатералям поступает в ретикулярную формацию. После многократных переключений в ее синапсах афферентное возбуждение достигает высших отделов головного мозга. Эти так называемые неспецифические активирующие влияния служат необходимым условием для поддержания бодрствования и осуществления любых поведенческих реакций. Помимо этого неспецифическая активация является важным условием для формирования селективных свойств нейронов коры в процессе онтогенетического созревания и обучения.
В аппарате восходящей ретикулярной формации сформировался механизм преобразования сенсорной информации в две формы активации: тоническую (генерализованную) и фазическую (локальную). Тоническая форма активации связана с функцией нижних стволовых отделов ретикулярной формации. Она генерализованно, диффузно поддерживает определенный уровень возбудимости в коре и подкорковых образованиях. Фазическая форма активации связана с верхними отделами ствола мозга, и прежде всего с неспецифической таламической системой, которая локально и избирательно распределяет воздействия восходящей активации на подкорковые образования, старую и новую кору.
Тоническая активация облегчается притоком возбуждений из различных органов чувств. «Экстренное» появление или исчезновение какого-либо раздражителя во внешней среде вызывает ориентировочный рефлекс и реакцию активации (экстренная мобилизация организма). Это поликомпонентная реакция, она связана с работой механизмов тонической и фазической активации ретикулярной формации (среднего мозга и неспецифических ядер таламуса). Кроме того, ориентировочный рефлекс связан с активирующей и тормозной функцией нейронов гиппокампа и хвостатого ядра, которые являются важным аппаратом регуляции тонических состояний коры мозга.
Кора головного мозга наряду со специфическим функциональным вкладом оказывает активирующие и тормозные влияния на нижележащие нервные образования. Корковые влияния, поступающие по нисходящим волокнам, представляют достаточно дифференцированную организацию и могут рассматриваться в качестве дополнительного третьего источника активации. Нисходящие волокна активирующей (и тормозящей) ретикулярной системы имеют достаточно дифференцированную корковую организацию; если связанные со специфическими путями пучки волокон (повышающих или понижающих тонус сенсорных или двигательных аппаратов) исходят из первичных (и частично из вторичных) зон коры, то волокна, опосредствующие более общие активирующие влияния на ретикулярную формацию ствола, исходят прежде всего из лобных отделов коры. Нисходящие волокна, идущие преимущественно от префронтальной (орбитальной и медиальной) коры, адресуются к ядрам зрительного бугра и нижележащих стволовых образований и являются тем аппаратом, посредством которого высшие отделы мозговой коры, непосредственно участвующие в формировании намерений и планов, управляют работой нижележащих аппаратов ретикулярной формации таламуса и ствола, тем самым модулируя их работу и обеспечивая наиболее сложные формы сознательной деятельности.
1.3 Блок программирования, запуска и контроля поведенческих актов
Прием, переработка и хранение внешней информации составляют только одну сторону психической жизни человека. Ее другую сторону составляет организация активной сознательной психической деятельности. С этой задачей и связан третий из основных функциональных блоков мозга - блок программирования, регуляции и контроля за протекающей деятельностью.
Аппараты третьего функционального блока мозга расположены в передних отделах больших полушарий, спереди от центральной извилины (см. рис. 1). Его основной отличительной чертой является то, что он не содержит модально-специфических зон, представляющих отдельные анализаторы, а состоит целиком из аппаратов эфферентного (двигательного) типа, однако сам находится под постоянным притоком информации из аппаратов афферентного (сенсорного) блока. Следующая важнейшая черта, отличающая работу третьего функционального блока от афферентного, состоит в том, что процессы здесь идут в нисходящем направлении, начинаясь с наиболее высоких - третичных и вторичных зон коры. Здесь в высших отделах интегративно-пускового блока формируются двигательные программы, а затем переходят к аппаратам низших моторных образований (первичным корковым зонам; стволовым и спинальным двигательным ядрам). Решающее значение в подготовке двигательной эфферентной импульсации имеют надстроенные над первичной моторной корой вторичные (премоторные отделы, 6-е и 8-е поля) и третичные зоны (префронтальные отделы лобной коры), или лобные доли (см. рис. 1).
Страницы: 1, 2
ИНТЕРЕСНОЕ
© 2009 Все права защищены. |