 | |
МЕНЮ
- Главная
- Языкознание филология
- Финансовые науки
- Управленческие науки
- Товароведение
- Технология
- Теплотехника
- Теория организации
- Теория государства и права
- Таможенная система
- Схемотехника
- Строительство
- Страхование
- Статистика
- Религия и мифология
- Психология и педагогика
- Промышленность производство
- Медицинские науки
- Медицина
- Краеведение и этнография
- Компьютерные науки
- История
- Искусство и культура
- Информатика
- Инвестиции
- Издательское дело и полиграфия
- Зоология
- Журналистика
- Естествознание
- Деньги и кредит
- Делопроизводство
- Гражданское право и процесс
- Государство и право
- Геополитика
- Геология
- Геодезия
- География
- Военная кафедра
- Ветеринария
- Валютные отношения
- Бухгалтерский учет и аудит
- Ботаника и сельское хоз-во
- Биржевое дело
- Биология и химия
- Биология
- Безопасность жизнедеятельности
- Банковское дело
- Астрономия
- Астрология
- Архитектура
- Арбитражный процесс
- Административное право
- Авиация и космонавтика
- Карта сайта
Контрольная работа: Механизмы реализации путей гибели клетки
Стволовые клетки способны восстанавливать кроветворение у облученных животных (радиозащитное действие), длительно поддерживать кроветворение и образовывать колониеобразующие единицы селезенки (двенадцатидневные селезеночные колонии), дающие начало гранулоцитарным, моноцитарным, эритроидным, мегакариоцитарным и лимфоидным колониям. Все клетки гемопоэтического происхождения образуются из примитивных стволовая кроветворная клеток (пСКК), локализованных в костном мозге и дающих начало клеткам четырех основных направлений дифференцировки:
мегакариоцитарного (тромбоциты),
миелоидного (гранулоциты и моноядерные фагоциты)
Дивергенция общего стволового элемента происходит на самом раннем этапе костномозговой дифференцировки.
Антигенпрезентирующие клетки в основном, но не исключительно, развиваются из миелоидных клеток-предшественников.
Клетки миелоидного и лимфоидного ряда наиболее важны для функционирования иммунной системы.
Лимфопоэтическая своловая клетка определяет две самостоятельные линии развития, приводящие к образованию Т-клеток и В-клеток.
Первая образующаяся из ГСК клетка-предшественник представляет собой колониеобразующуюся единицу (КОЕ) , которая определяет линии развития, приводящие к образованию гранулоцитов, эритроцитов, моноцитов и мегакариоцитов. Созревание этих клеток происходит под влиянием колониестимулирующих факторов (КСФ) и ряда интерлейкинов, в том числе ИЛ-1, ИЛ-3, ИЛ-4, ИЛ-5 и ИЛ-6. Все они играют важную роль в положительной регуляции (стимуляции) гемопоэза и продуцируются, главным образом, стромальными клетками костного мозга, но также и зрелыми формами дифференцированных миелоидных и лимфоидных клеток. Другие цитокины (например, ТРФ-бета) могут осуществлять понижающую регуляцию (подавление) гемопоэза).
У всех клеток как лимфоидного, так и миелоидного ряда время жизни ограничено, и все они непрерывно образуются.
У млекопитающих в период внутриутробного развития ГСК присутствуют в желточном мешке, печени, селезенке и костном мозге. Во взрослом организме гемопоэтические стволовые клетки находятся в основном в костном мозге, где они в норме довольно редко делятся, производя новые стволовые клетки (самообновление). Животное можно спасти от последствий облучения в летальных дозах введением клеток костного мозга, которые заселяют его лимфоидную и миелоидную ткани.
Плюрипотентные стволовые клетки дают начало коммитированным клеткам-предшественницам, которые уже необратимо определились как предки кровяных клеток одного или нескольких типов. Полагают, что коммитированные клетки делятся быстро, но ограниченное число раз, при этом делятся они под воздействием факторов микроокружения: соседних клеток и растворимых или мембраносвязанных цитокинов. В конце такой серии делений клетки эти становятся терминально дифференцированными, обычно больше не делятся и погибают через несколько дней или недель. Плюрипотентные стволовые клетки малочисленны, их трудно распознавать, и все еще неясно, как они выбирают свой путь среди разных вариантов развития. Программирование клеточных делений и выведение клеток на определенный путь дифференцировки (коммитирование), видимо, включают в себя и случайные события. Стволовая клетка плюрипотентна, т.к. дает начало многим видам терминально дифференцированных клеток. Что касается клеток крови, то эксперименты показывают, что все классы клеток крови - и миелоидных и лимфоидных - происходят от общей гемопоэтической стволовой клетки.
Гемопоэтическая стволовая клетка развивается следующим образом. У эмбриона гемопоэз начинается в желточном мешке, но по мере развития эта функция переходит к печени плода и, наконец, к костному мозгу, где и продолжается в течение всей жизни. Гемопоэтическая стволовая клетка, дающая начало всем элементам крови, плюрипотентна и заселяет другие гемо - и лимфопоэтические органы и самовоспроизводится, превращаясь в новые стволовые клетки. Животное можно спасти от последствий облучения в летальных дозах введением клеток костного мозга, которые заселяют его лимфоидную и миелоидную ткани.
Во взрослом организме гемопоэтические стволовые клетки находятся главным образом в костном мозге, где они в норме довольно редко делятся, производя новые стволовые клетки (самообновление).
Клетку-предшественницу, дающую в культуре клеток начало колонии эритроцитов, называют колониеобразующей единицей эритроидного ряда, или КОЕ-Э, и она дает начало зрелым эритроцитам после шести или даже меньшего числа циклов деления. КОЕ-Э еще не содержит гемоглобин.
Гемопоэзом (haemopoesis) называют развитие крови. Различают эмбриональный гемопоэз, который происходит в эмбриональный период
и приводит к развитию крови как ткани, и постэмбриональный гемопоэз, который представляет собой процесс физиологической регенерации крови. Развитие эритроцитов называют эритропоэзом, развитие гранулоцитов - гранулоцитопоэзом, тромбоцитов - тромбоцитопоэзом, развитие моноцитов - моноцитопоэзом, развитие лимфоцитов и иммуноцитов - лимфоцито - и иммуноцитопоэзом.
Эмбриональный гемопоэз.
В развитии крови как ткани в эмбриональный период можно выделить 3 основных этапа, последовательно сменяющих друг друга:
1) мезобластический, когда начинается развитие клеток крови во внезародышевых органах - мезенхиме стенки желточного мешка, хориона и стебля (с 3-й по 9-ю неделю развития зародыша человека) и появляется первая генерация стволовых клеток крови (СКК);
2) печеночный, который начинается в печени с 5-6-й недели развития плода, когда печень становится основным органом гемопоэза, в ней образуется вторая генерация СКК.
Кроветворение в печени достигает максимума через 5 месяцев и завершается перед рождением. СКК печени заселяют тимус (здесь, начиная с 7-8-й недели, развиваются Т-лимфоциты), селезенку (гемопоэз начинается с 12-й недели) и лимфатические узлы (гемопоэз отмечается с 10-й недели);
3) медуллярный (костномозговой) - появление третьей генерации СКК в костном мозге, где гемопоэз начинается с 10-й недели и постепенно нарастает к рождению, а после рождения костный мозг становится центральным органом гемопоэза.
Кроветворение в стенке желточного мешка. У человека оно начинается в конце 2-й - начале 3-й недели эмбрионального развития. В мезенхиме стенки желточного мешка обособляются зачатки сосудистой крови, или кровяные островки. В них мезенхимные клетки округляются, теряют отростки и преобразуются в стволовые клетки крови. Клетки, ограничивающие кровяные островки, уплощаются, соединяются между собой и образуют эндотелиальную выстилку будущего сосуда. Часть СКК дифференцируется в первичные клетки крови (бласты), крупные клетки с базофильной цитоплазмой и ядром, в котором хорошо заметны крупные ядрышки. Большинство первичных кровяных клеток митотически делится и превращается в первичные эритробласты, характеризующиеся крупным размером (мегалобласты). Это превращение совершается в связи с накоплением эмбрионального гемоглобина в цитоплазме бластов, при этом сначала образуются полихроматофильные эритробласты, а затем оксифилъные эритробласты с большим содержанием гемоглобина. В некоторых первичных эритробластах ядро подвергается кариорексису и удаляется из клеток, в других ядро сохраняется. В результате образуются безъядерные и ядросодержащие первичные эритроциты, отличающиеся большим размером по сравнению с нормоцитами и поэтому получившие название мегалоцитов. Такой тип кроветворения называется мегалобластическим. Он характерен для эмбрионального периода, но может появляться в постнатальном периоде при некоторых заболеваниях (злокачественное малокровие). Наряду с мегалобластическим в стенке желточного мешка начинается нормобластическое кроветворение, при котором из бластов образуются вторичные эритробласты; сначала они превращаются в полихроматофильные эритробласты, далее в нормобласты, из которых образуются вторичные эритроциты (нормоциты); размеры последних соответствуют эритроцитам (нормоцитам) взрослого человека. Развитие эритроцитов в стенке желточного мешка происходит внутри первичных кровеносных сосудов, т.е. интраваскулярно. Одновременно экстраваскулярно из бластов, расположенных вокруг сосудистых стенок, дифференцируется небольшое количество гранулоцитов - нейтрофилов и эозинофилов. Часть СКК остается в недифференцированном состоянии и разносится током крови по различным органам зародыша, где происходит их дальнейшая дифференцировка в клетки крови или соединительной ткани. После редукции желточного мешка основным кроветворным органом временно становится печень.
Кроветворение в печени. Печень закладывается примерно на 3-4-й неделе эмбриональной жизни, а с 5-й недели она становится центром кроветворения. Кроветворение в печени происходит экстраваскулярно, по ходу капилляров, врастающих вместе с мезенхимой внутрь печеночных долек. Источником кроветворения в печени являются стволовые клетки крови, из которых образуются бласты, дифференцирующиеся во вторичные эритроциты. Процесс их образования повторяет описанные выше этапы образования вторичных эритроцитов. Одновременно с развитием эритроцитов в печени образуются зернистые лейкоциты, главным образом нейтрофильные и эозинофильные. В цитоплазме бласта, становящейся более светлой и менее базофильной, появляется специфическая зернистость, после чего ядро приобретает неправильную форму. Кроме гранулоцитов, в печени формируются гигантские клетки - мегакариоциты. К концу внутриутробного периода кроветворение в печени прекращается.
Кроветворение в тимусе. Тимус закладывается в конце 1-го месяца внутриутробного развития, и на 1-8-й неделе его эпителий начинает заселяться стволовыми клетками крови, которые дифференцируются в лимфоциты тимуса. Увеличивающееся число лимфоцитов тимуса дает начало Т-лимфоцитам, заселяющим Т-зоны периферических органов иммунопоэза.
Кроветворение в селезенке. Закладка селезенки происходит в конце 1-го месяца эмбриогенеза. Из вселяющихся сюда стволовых клеток происходит экстраваскулярное образование всех видов форменных элементов крови, т.е. селезенка в эмбриональном периоде представляет собой универсальный кроветворный орган. Образование эритроцитов и гранулоцитов в селезенке достигает максимума на 5-м месяце эмбриогенеза. После этого в ней начинает преобладать лимфоцитопоэз.
Кроветворение в лимфатических узлах. Первые закладки лимфатических узлов человека появляются на 7-8-й неделе эмбрионального развития. Большинство лимфатических узлов развивается на 9-10-й неделе. В этот же период начинается проникновение в лимфатические узлы стволовых клеток крови, из которых на ранних стадиях дифференцируются эритроциты, гранулоциты и мегакариоциты. Однако формирование этих элементов быстро подавляется образованием лимфоцитов, составляющих основную часть лимфатических узлов. Появление единичных лимфоцитов происходит уже в течение 8-15-й недели развития, однако массовое "заселение" лимфатических узлов предшественниками Т - и В-лимфоцитов начинается с 16-й недели, когда формируются посткапиллярные венулы, через стенку которых осуществляется процесс миграции клеток. Из клеток-предшественников дифференцируются лимфобласты (большие лимфоциты), а далее средние и малые лимфоциты. Дифференцировка Т - и В-лимфоцитов происходит в Т - и В-зависимых зонах лимфатических узлов.
Кроветворение в костном мозге. Закладка костного мозга осуществляется на 2-м месяце эмбрионального развития. Первые гемопоэтические элементы появляются на 12-й неделе развития; в это время основную массу их составляют эритробласты и предшественники гранулоцитов. Из СКК в костном мозге формируются все форменные элементы крови, развитие которых происходит экстраваскулярно. Часть СКК сохраняется в костном мозге в недифференцированном состоянии, они могут расселяться по другим органам и тканям и являться источником развития клеток крови и соединительной ткани. Таким образом, костный мозг становится центральным органом, осуществляющим универсальный гемопоэз, и остается им в течение постнатальной жизни. Он обеспечивает стволовыми кроветворными клетками тимус и другие гемопоэтические органы.
Постэмбриональный гемопоэз. Постэмбриональный гемопоэз представляет собой процесс физиологической регенерации крови (клеточное обновление), который компенсирует физиологическое разрушение дифференцированных клеток.
Миелопоэз происходит в миелоидной ткани (textus myeloideus), расположенной в эпифизах трубчатых и полостях многих губчатых костей.
Здесь развиваются форменные элементы крови: эритроциты, гранулоциты, моноциты, кровяные пластинки, предшественники лимфоцитов.
В миелоидной ткани находятся стволовые клетки крови и соединительной ткани.
Предшественники лимфоцитов постепенно мигрируют и заселяют такие органы, как тимус, селезенка, лимфатические узлы и др.
Лимфопоэз происходит в лимфоидной ткани (textus lymphoideus), которая имеет несколько разновидностей, представленных в тимусе, селезенке, лимфатических узлах. Она выполняет основные функции: образование Т - и В-лимфоцитов и иммуноцитов (плазмоцитов и др.).
Миелоидная и лимфоидная ткани являются разновидностями соединительной ткани, т.е. относятся к тканям внутренней среды. В них представлены две основные клеточные линии - клетки ретикулярной ткани и гемопоэтические.
Ретикулярные, а также жировые, тучные и остеогенные клетки вместе с межклеточным веществом (матрикс) формируют микроокружение для
гемопоэтических элементов. Структуры микроокружения и гемопоэтические
клетки функционируют в неразрывной связи. Микроокружение оказывает
воздействие на дифференцировку клеток крови (при контакте с их рецепторами или путем выделения специфических факторов).
Для миелоидной и всех разновидностей лимфоидной ткани характерно
наличие стромальных ретикулярных и гемопоэтических элементов,
образующих единое функциональное целое. В тимусе имеется сложная строма, представленная как соединительнотканными, так и ретикулоэпителиальными клетками. Эпителиальные клетки секретируют особые вещества - тимозины, оказывающие влияние на дифференцировку из СКК Т-лимфоцитов. В лимфатических узлах и селезенке специализированные ретикулярные клетки создают микроокружение, необходимое для пролиферации и дифференцировки в специальных Т - и В-зонах Т - и В-лимфоцитов и плазмоцитов.
СКК являются плюрипотентными (полипотентными) предшественниками всех клеток крови и относятся к самоподдерживающейся популяции клеток. Они редко делятся. Впервые представление о родоначальных клетках крови сформулировал в начале XX в.А. А. Максимов, который считал, что по своей морфологии они сходны с лимфоцитами. В настоящее время это представление нашло подтверждение и дальнейшее развитие в новейших экспериментальных исследованиях, проводимых главным образом на мышах. Выявление СКК стало возможным при применении метода колониеобразования.
Экспериментально (на мышах) показано, что при введении смертельно облученным животным (утратившим собственные кроветворные клетки) взвеси клеток красного костного мозга или фракции, обогащенной СКК, в селезенке появляются колонии клеток - потомков одной СКК. Пролиферативную активность СКК модулируют колониестимулирующие факторы (КСФ), интерлейкины (ИЛ-3 и др.). Каждая СКК в селезенке образует одну колонию и называется селезеночной колониеобразующей единицей (КОЕ-С).
Подсчет колоний позволяет судить о количестве стволовых клеток, находящихся во введенной взвеси клеток. Таким образом, было установлено, что у мышей на 105 клеток костного мозга приходится около 50 стволовых клеток, из селезенки - 3,5 клетки, среди лейкоцитов крови - 1,4 клетки.
Исследование очищенной фракции стволовых клеток с помощью электронного микроскопа позволяет считать, что по ультраструктуре они очень близки к малым темным лимфоцитам.
Исследование клеточного состава колоний позволило выявить две линии их дифференцировки. Одна линия дает начало мультипотентной клетке - родоначальнице гранулоцитарного, эритроцитарного, моноцитарного и мегакариоцитарного рядов гемопоэза (КОЕ-ГЭММ). Вторая линия дает начало мультипотентной клетке - родоначальнице лимфопоэза (КОЕ-Л). Из мультипотентных клеток дифференцируются олигопотентные (КОЕ-ГМ) и унипотентные родоначальные (прогениторные) клетки.
Методом колониеобразования определены родоначальные унипотентные клетки для моноцитов (КОЕ-М), нейтрофильных гранулоцитов (КОЕ-Гн), эозинофилов (КОЕ-Эо), базофилов (КОЕ-Б), эритроцитов (БОЕ-Э и КОЕ-Э), мегакариоцитов (КОЕ-МГЦ), из которых образуются клетки-предшественники (прекурсорные). В лимфопоэтическом ряду выделяют унипотентные клетки - предшественницы для В-лимфоцитов и соответственно для Т - лимфоцитов. Полипотентные (плюрипотентные и мультипотентные), олигопотентные и унипотентные клетки морфологически не различаются.
Все приведенные выше стадии развития клеток составляют четыре основных компартмена: I - стволовые клетки крови (плюрипотентные, полипоте нтные); II - коммитированные родоначальные клетки (мультипотентные); III - коммитированные родоначальные (прогенторные) олигопотентные и унипотентные клетки; IV - клетки-предшественники (прекурсорные).
Дифференцировка полипотентных клеток в унипотентные определяется действием ряда специфических факторов - эритропоэтинов (для эритробластов), гранулопоэтинов (для миелобластов), лимфопоэтинов (для лимфобластов), тромбопоэтинов (для мегакариобластов) и др.
Из каждой клетки-предшественницы происходит образование конкретного вида клеток. Созревание каждого вида клеток проходит ряд стадий, которые в совокупности образуют компартмент созревающих клеток (V).
Зрелые клетки представляют последний компартмент (VI). Все клетки V и VI компартментов морфологически можно идентифицировать.
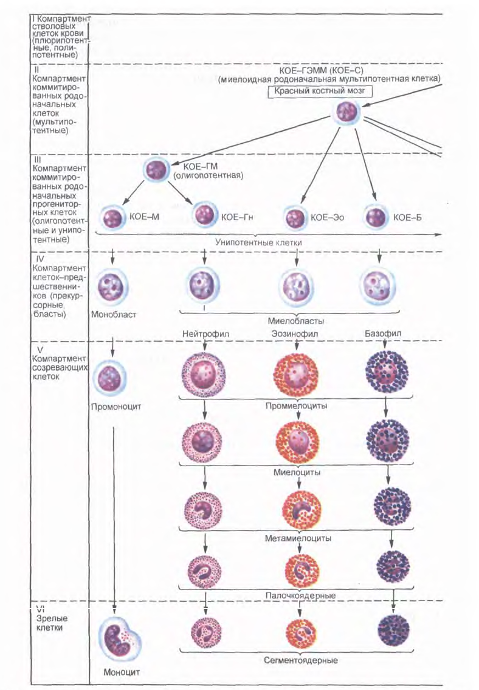
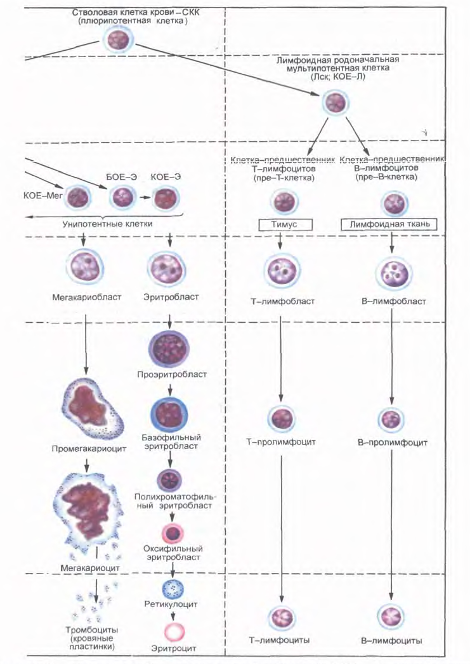
Рис.18. Постэмбриональный гемопоэз, окраска азур 11-эозином (схема по НАЮриной). Стадии дифферениировки крови: I-IV - морфологически неидентифицируемые клетки; V - VI - морфологически идентифицируемые клетки. Б - базофил; БОЕ - бурстобраэуюшая единица; Г - гранулоциты; Гн - гранулоцит нейтрофильный; КОЕ - колониеобразующ! единицы; КОЕ-С - селезеночная колониеобразующая единица; Л - лимфоцит; Лек - mt фоидная стволовая клетка; М - моноцит; Мет - мегакариошгг; Эо - эозинофил; Э - эритроцит.
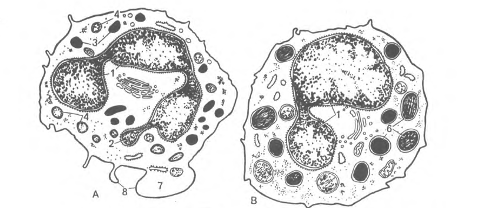
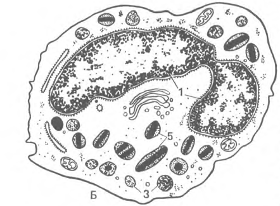
Рис. 19. Ультрамикроскопическое строение гранулоцитов (схема по НА. Юриной и Л.С. Румянцевой).
А - сегментоядерный нейтрофильный гранулоцит; Б – эозинофильный (ацидофильный) гранулоиит; В - базофильный фанулоцит: 1 - сегменты ядра; 2 - тельце полового хроматина; 3 – первичные (азурофильные) гранулоциты; 4 - вторичные (специфические) гранулы; 5 - зрелые специфические гранулы эозинофила, содержащие кристаллоиды; б - гранулы базофила различной величины и плотности; 7 - периферическая зона, не содержащая органелл; 8 – микроворсинки и псевдоподии.
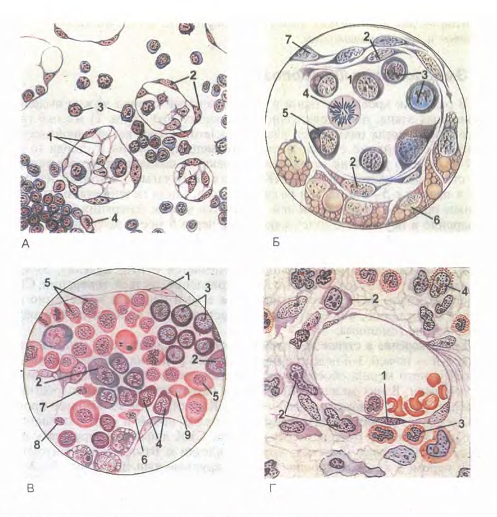
Рис. 20. Эмбриональный гемоппэп (по А.А. Максимову).
А - кроветворение в стенке желточного мешка зародыша морской свинки: 1 - меэенхималыгые клетки; 2 - эндотелий стенки сосудов; 3 - первичные кровяные клетки-бласты; 4 - митотическос деление бластов; Б - поперечный срез кровяного островка зародыша кролика S'/j сут: I - полость сосуда; 2 - эндотелий; 3 - интраваскулярные кровяные клетки; 4 - делящаяся кровяная клетка; 5 - формирование первичной кровяной клетки; 6 - энтодерма; 7 - висцеральный листок мезодермы. В - развитие вторичны); эритробластов в сосуде зародыша кролика 13'Д сут: 1 - эндотелий; 2 - проэритробласты; 3 - базофильные эритробласты; 4 - полихроматофильные эритробласты; 5 - оксифильные эритробласты (нормобласты); 6 - оксифильный эритробласт с пикнотическим ядром; 7 - обособление ядра от оксифильного эритробласта (нормобласта); 8 - вытолкнутое ядро нормобласта; 9 - вторичный эритроцит. Г - кроветворение в костном мозге зародыша человека с длиной тела 77 мм. Экстра во скул я рное развитие клеток крови: 1 - эндотелий сосуда; 2 - бласты; 3 - нейтрофильные гранулоциты; 4 - эоэинофильный миелоцит.
Механизмы реализации путей гибели клеток.